





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
夜晚蓝光和白光暴露对肝脏生物钟节律和肝脏炎症反应影响的差异
[梁昕悦1, 2  , 曾晶
, 曾晶2 , 艾常鹏3 , 罗杰承3 , 范晓静4 , 吴胜男3, * , 顾瑛1, 2, 3, * ]
, 曾晶, 顾瑛]
|
|


作者简介: 梁昕悦, 女, 1993年生, 解放军医学院博士研究生 e-mail: 472465726@qq.com
光对昼夜节律的调控属于非成像视觉功能, 以自主感光视神经节细胞为主的非成像视觉效应细胞对机体的生物节律、 代谢和认知等有显著影响, 主要由蓝光介导。 该研究探索夜晚蓝光和白光暴露对肝脏生物钟节律以及肝脏炎症反应的影响。 将沙鼠每晚暴露在蓝光或白光中1或3 h, 持续10周, 采用实时荧光定量PCR检测肝脏生物钟基因表达节律, 采用转录组学分析肝脏全部基因表达水平, 采用H&E染色和免疫荧光法观察肝脏组织形态及炎症因子水平。 研究结果显示, 夜晚光照对肝脏多种生物钟基因 Arntl、 Clock、 Cry1、 Nr1 d1、 Per2和 Ror-α表达节律均造成显著破坏。 夜晚光照对肝脏免疫系统基因表达的影响最显著。 夜晚光照可引起肝细胞水肿, 上调促炎因子白介素17A和粒细胞-巨噬细胞集落刺激因子的表达。 夜晚光照暴露扰乱了肝脏生物钟节律并引发肝脏炎症反应。 该研究凸显了非成像视觉通路在肝脏代谢稳态中的关键作用。
, ZENG Jing, GU Ying
The regulation of circadian rhythm by light is a non-imaging visual function.Non-imaging visual effect cells, mainly intrinsically photosensitive retinal ganglion cells, significantly impact the body's biological rhythm, metabolism, cognition, etc., primarilymediated by blue light. This study aimed to explore the effects of night light exposure on liver circadian rhythm and inflammation. The gerbils were exposed to white and blue light for 1 or 3 hours every night for 10 weeks. The real-time reverse transcription-polymerase chain reaction was used to detect the expression rhythm of liver clock genes; transcriptomics was used to analyze the expression level of all liver genes; hematoxylin-eosin staining was used to detect liver tissue morphology, and immunofluorescence staining was used to detect the level of inflammatory factors in liver tissue. The results showed that light exposure at night significantly disrupted the liver's expression rhythm of Arntl, Clock, Cry1, Nr1d1, Per2, and Rora. Light exposure at night had the most significant effect on liver immune system gene expression. Light exposure at night induced hepatocyte edema and up-regulated the expression of pro-inflammatory factors interleukin 17A and granulocyte-macrophage colony-stimulating factor. In short, exposure to light at night disrupted liver circadian rhythms and triggered an inflammatory response in the liver. This study highlights the critical role of non-imaging visual pathways in hepatic metabolic homeostasis.
昼夜节律是生物为适应地球24 h自转而进化出的周期性生命活动, 从细菌、 植物、 无脊椎动物到哺乳动物, 几乎所有生物的生理活动均有昼夜节律变化, 比如睡眠-觉醒周期、 激素的分泌、 体温和血压等[1]。 昼夜节律由内源生物钟控制, 可在没有外界信号刺激下自主振荡。 生物钟本质上是构成自主振荡转录— 翻译负反馈环路的一组基因。 其核心正向分子是大脑/肌肉芳香烃受体核转位因子样蛋白1(brain and muscle ARNT-like1, BMAL 1)和昼夜运动输出周期蛋白失效(circadian locomotor output cycles protein kaput, CLOCK), 负向分子是周期蛋白(period, PER)和隐色素(cryptochrome, CRY)。 视黄酸受体相关孤儿受体(retinoic acid receptor-related orphan receptor, ROR)和孤儿核受体(nuclear receptor subfamily 1 group D member 1, REV-ERBα )构成哺乳动物时钟分子机制的调节旁路[2]。
生理状态下, 生物钟受周期性变化的环境参数影响, 比如光照、 温度、 湿度、 食物和社交活动, 使生物钟的周期与外界环境参数的变化周期保持一致, 维持同步[3], 光是生物钟与外界环境同步最重要的信号[4]。 光信号主要通过“ 视网膜-下丘脑通路” 调节昼夜节律[5]。 昼夜节律的光感受器是一种含有视黑蛋白的自主感光视神经节细胞[6], 对波长480 nm的蓝光最敏感, 能够将光信号转化为电信号, 投射到视交叉上核等参与非成像视觉功能的脑区, 通过神经元和内分泌信号同步外周生物钟, 参与非成像视觉功能的调节[7]。 视杆细胞和视锥细胞也参与非成像视觉效应的调节[8]。
人工光的使用打破了自然光光谱、 强度和时间的节律性变化, 导致昼夜节律紊乱。 代谢是生命最基本的生理活动, 广泛受到生物钟的调节。 目前光谱对代谢影响的研究结论尚不统一, 有研究发现晚上9~10点暴露在蓝光下1 h影响沙鼠葡萄糖耐量[9]。 也有研究发现小鼠暴露在蓝光下8周, 白色脂肪相关生物钟基因Clock、 Arntl、 Per1、 Cry1、 Cry2、 Rorα 、 Rev-erbα 和Rev-erbβ 表达上调, 小鼠出现脂质代谢障碍和脂肪细胞肥大[10]。 Zhang等[11]将小鼠在150 lux不同波长的正常光周期下暴露8周, 研究发现与白光暴露相比, 绿光暴露的小鼠体重明显增加, 同时出现糖耐量下降和脂代谢异常, 红光暴露小鼠也显示出轻度糖耐量受损, 但蓝光暴露小鼠并未表现出现明显的糖脂代谢异常。
21世纪初, 人口结构逐渐由农村转向城市, 目前有44亿人居住在城市, 城市化的转变意味着大多数人在晚上暴露在更高水平的光线下, 夜间照明正在全球范围内增加[12]。 2014年美国疾病控制与预防中心的研究调查数据显示, 随着人工光污染程度的增加, 代谢紊乱的流行呈指数级增长[13]。 通过卫星遥感获得室外环境夜间光发射的图像, 发现夜间暴露在室外富含蓝光的光线下, 会增加患乳腺癌、 前列腺癌和结肠癌的风险[14]。
本研究将沙鼠每晚暴露在蓝光或白光中1或3 h, 持续10周, 探索夜晚光照对肝脏生物钟节律以及肝脏炎症的影响。 为非成像视觉通路在肝脏代谢稳态中的作用提供一定参考。
本研究选用八周龄雄性长爪沙鼠(Meriones unguiculatus)作为实验动物, 长爪沙鼠为日行性啮齿类动物, 为小型草原动物, 主要分布于我国的内蒙古自治区及其毗邻的省区, 又名蒙古沙鼠, 清洁级封闭群, 英文品系缩写为CMU, 购买自斯贝福(北京)生物技术有限公司。 沙鼠视网膜含有的感光细胞相对夜行性小鼠较多, 更接近于人类的视网膜结构[15], 同时考虑到人是日行性动物, 实验结果更具有参考意义。 沙鼠均饲养在实验室自主研制的昼夜节律实验柜中, 实验柜内顶部装有发光二极管光板, 由计算机控制照明条件, 可根据不同分组控制照度(luminous, lux)、 色温(kelvin, K)及开关灯时间(zeitgeber time, ZT)。 实验柜恒温恒湿通风, 环境温度控制在(21± 1) ℃, 湿度控制在50%~60%, 每个柜子放置一个鼠笼, 每个鼠笼饲养2只沙鼠, 沙鼠可自由活动、 进食和饮水。 饲养环境为12 h明∶ 12 h暗, 色温为4 000 K中性白光。 照度依据前期研究成果, 模拟了人类日间与夜间的光照强度保持一致, 均采用78 lux光强。 夜晚开灯时间选择在ZT14, 由于既往研究发现夜晚在ZT14开灯1 h后严重损害沙鼠葡萄糖耐量[9], 同时也为了更好的模拟夜晚加班状态。 ZT0表示开灯时间。
实验前, 沙鼠在12 h明∶ 12 h暗的环境中, 给予正常饮食适应性饲养2周, 照度78 lux。 实验开始后, 将沙鼠分为对照组和夜晚光照组, 其中对照组即饲养环境光照条件, 而夜晚光照组在ZT14接受白光(4 000 K, 78 lux)或蓝光(471 nm, 78 lux)照明1或3 h, 随机分成5组, 每组12~14只, 持续10周。 实验结束后, 沙鼠禁食过夜, 分别在ZT1、 ZT7、 ZT13和ZT19处死并收集肝脏组织。 实验期间所有动物操作均符合动物福利伦理。
采用异丙醇沉淀法抽提总RNA, 反转录合成cDNA, 采用实时荧光定量PCR技术检测生物钟基因Arntl(编码BMAL1蛋白)、 Clock(编码Clock蛋白)、 Per2(编码Per2蛋白)、 Cry1(编码CRY1蛋白)、 Rora(编码ROR-α 蛋白)和Nr1d1(编码REV-ERBα 蛋白)在肝脏组织中的表达。 引物信息见表1。
| 表1 RT-PCR引物序列 Table 1 Primer sequence for RT-PCR |
TRIzol试剂提取总RNA。 样品送至上海美吉生物医药科技有限公司进行转录组测序分析。 具体步骤包括转录组测序数据的质量评估、 与参考基因组的比较分析、 基因表达水平分析、 表达差异分析以及KEGG分析。
取部分肝脏组织并将其储存于4%多聚甲醛固定液中。 石蜡切片制备、 H& E染色、 免疫荧光染色和图像采集由武汉赛维尔生物科技有限公司完成。 使用NIKON ECLIPSE E100立式光学显微镜获取H& E图像。 用于免疫荧光染色的抗体信息: 白细胞介素17A, 抗体货号为ab214588(abcam), 二抗使用Alexa Fluor 488标记山羊抗兔IgG, 检测光谱为激发波长465~495 nm, 发射波长515~555 nm。 粒细胞-巨噬细胞集落刺激因子, 抗体货号为pa5-79075(thermo), 二抗使用Cy3标记山羊抗兔IgG, 检测光谱为激发波长510~560 nm, 发射波长590 nm。 4’ , 6-二脒基-2-苯基吲哚的检测光谱为激发波长330~380 nm, 发射波长420 nm。 使用NIKON ECLIPSE C1立式荧光显微镜捕获荧光图像。
采用SPSS 25.0统计学软件进行数据分析, Graphpad Prism 10软件展示数据, 统计学结果采用平均值± 标准差表示。 其中两组样本组间比较采用t检验, 每种实验均至少重复3次, * p< 0.05为有统计学差异。
图1(a, b, c)所示, 为了探索夜晚光照对沙鼠肝脏生物钟基因表达节律的影响, 将沙鼠每晚暴露在白光或蓝光中1或3 h。 图2所示, 结果发现夜晚光照引起肝脏组织生物钟基因Arntl、 Clock、 Cry1、 Nr1d1、 Per2、 Ror-α 表达节律的不同程度破坏。 整体来讲, 与暴露1 h相比, 暴露3 h对生物钟基因表达节律影响比较显著, 表现为对基因节律的几乎完全逆转、 过低或过高表达。 与照射白光相比, 照射蓝光对生物钟基因表达节律的破坏更显著。 表明夜晚光照破坏肝脏生物钟基因表达节律, 并且夜晚蓝光及长时间光照危害更大。
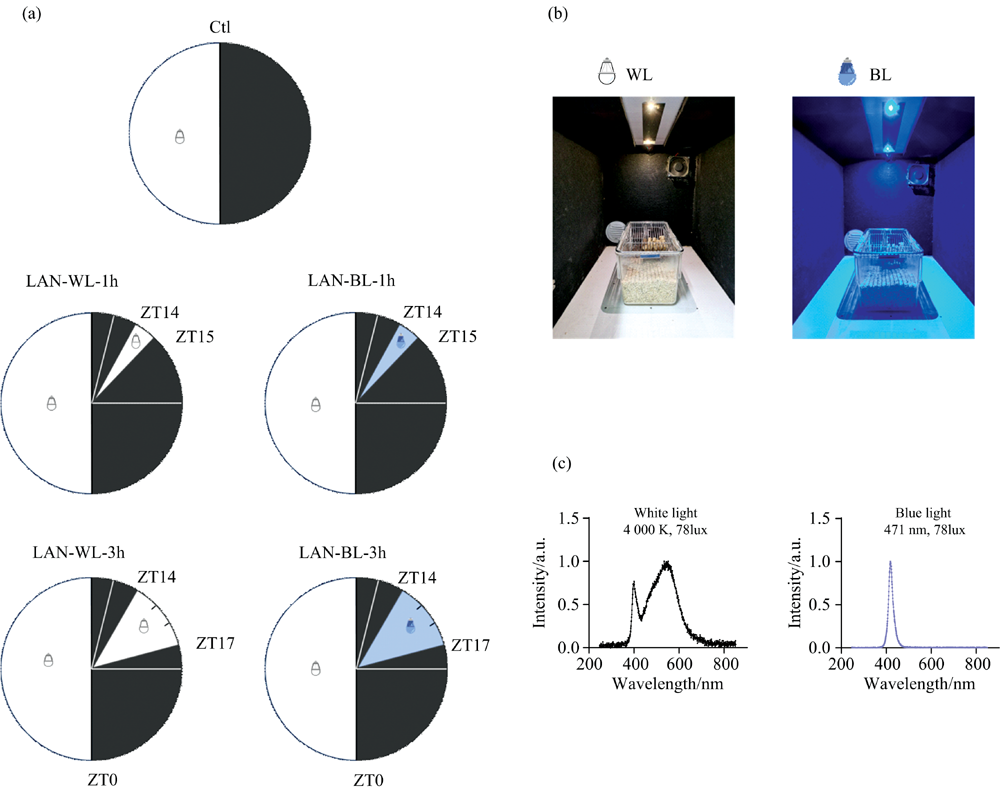 | 图1 夜晚光照设置 (a): 实验分组; (b): 照明环境; (c): 光谱 注: Ctl: 对照组; LAN-WL-1h: 夜晚白光1 h组; LAN-WL-3h: 夜晚白光3 h组; LAN-BL-1h: 夜晚蓝光1 h组; LAN-BL-3h: 夜晚蓝光3 h组; ZT: 授时因子时间, ZT0即开灯时间, ZT12即关灯时间; WL: 白光; BL: 蓝光; K: 开尔文; lux: 勒克斯Fig.1 Lighting settings (a): LAN diagram; (b): Lighting environment; (c): Spectrum Note: Ctl: Control group; LAN-WL-1h: White light exposure at night for 1 hour group; LAN-WL-3h: White light exposure at night for 3 hours group; LAN-BL-1h: Blue light exposure at night for 1 hour group; LAN-BL-3h: Blue light exposure at night for 3 hours group; ZT: Zeitgeber time, ZT0 is the time when lights are turned on, ZT12 is the time when lights are turned off; WL: White light; BL: Blue light; K: Kelvin; lux: Luminance |
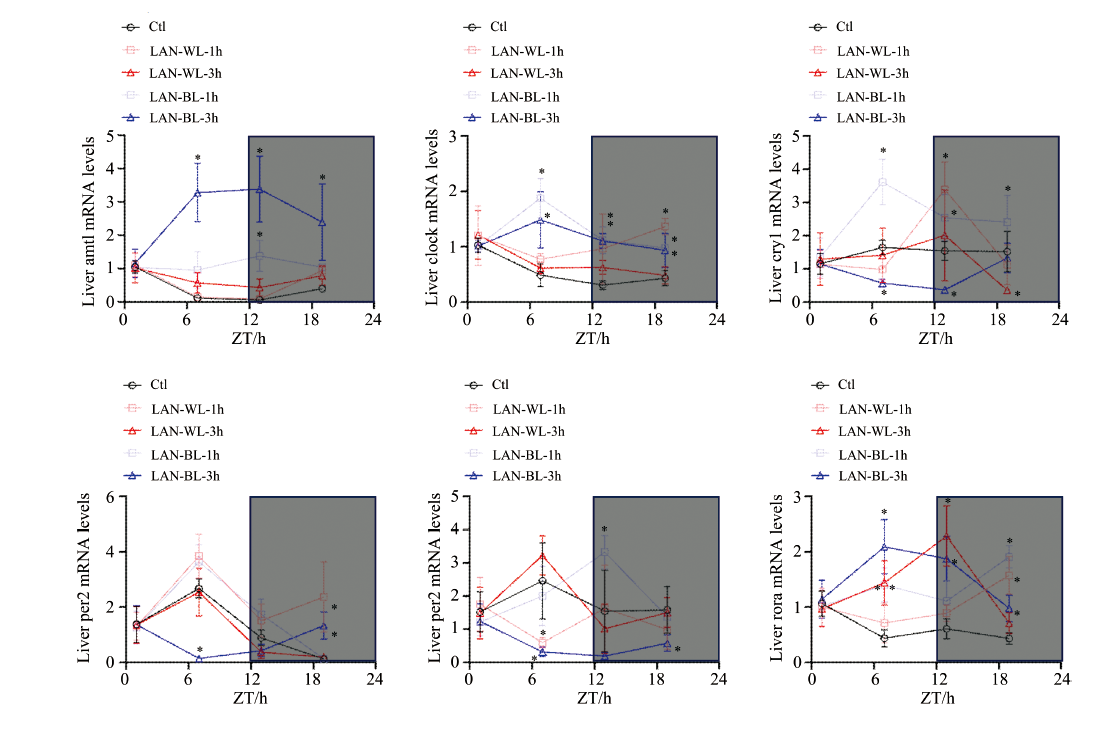 | 图2 夜晚光照对沙鼠肝脏组织生物钟基因Arntl、 Clock、 Cry1、 Nr1d1、 Per2、 Rorα 表达节律的影响(n=3; * p< 0.05 vs. 对照组) 注: Ctl: 对照组; LAN-WL-1h: 夜晚白光1 h组; LAN-WL-3h: 夜晚白光3 h组; LAN-BL-1h: 夜晚蓝光1 h组; LAN-BL-3h: 夜晚蓝光3 h组; ZT: 授时因子时间, ZT0即开灯时间, ZT12即关灯时间; Arntl: 芳香烃受体核转位因子样蛋白1; Clock: 昼夜运动输出周期蛋白失效; Cry1: 隐花色素1; Per2: 周期蛋白2; Nr1d1: 核受体亚家族1D组成员1; Rorα : 视黄酸受体相关孤儿受体基因α Fig.2 The impact of nighttime illumination on the rhythmic expression of circadian clock genes Arntl, Clock, Cry1, Nr1d1, Per2 and Rorα in gerbil liver tissue Note: Ctl: Control group; LAN-WL-1h: White light exposure at night for 1 hour group; LAN-WL-3h: White light exposure at night for 3 hours group; LAN-BL-1h: Blue light exposure at night for 1 hour group; LAN-BL-3h: Blue light exposure at night for 3 hours group; ZT: Zeitgeber time, ZT0 is the time when lights are turned on, ZT12 is the time when lights are turned off; Arntl: Aryl hydrocarbon receptor nuclear translocator like; Clock: Circadian locomator output cycles pretein kaput; Cry1: Cryptochrome 1; Per2: Period 2; Nr1d1: Nuclear receptor subfamily 1 group D member 1; Rorα : Retinoid-related orphan receptor alpha |
图3所示, KEGG功能注释分析显示, 差异表达的基因主要参与了代谢过程、 环境信息处理、 细胞过程、 生物体系统和人类疾病等内在生物学过程。 其中, 夜晚光照对外源物质生物降解和代谢的影响显著, 主要影响免疫系统以及内分泌系统, 提示可能引发感染性疾病、 癌症、 免疫疾病和心血管疾病。 图4所示, KEGG功能富集分析显示, 差异表达的基因主要集中在核因子κ B信号通路, 参与细胞吞噬, 造血干细胞分化, 天然免疫信号通路和获得性免疫信号通路, 提示可能引起原发性免疫缺陷等免疫系统疾病。
 | 图3 KEGG功能注释分析Fig.3 Analysis of KEGG function annotation |
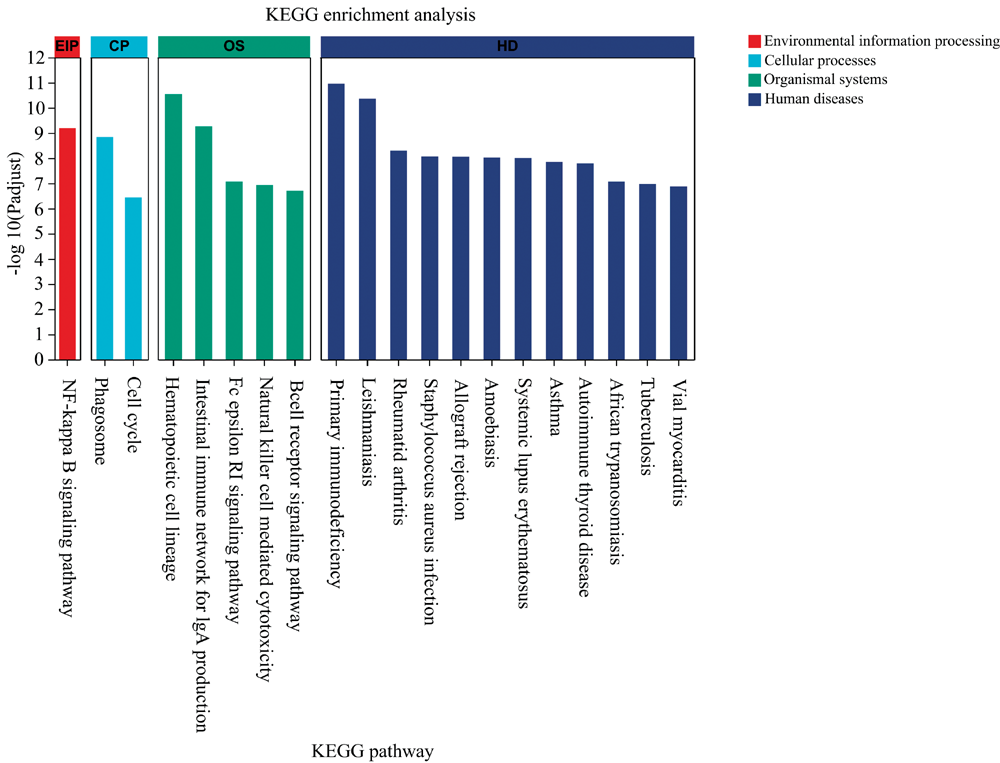 | 图4 KEGG功能富集分析Fig.4 KEGG functional enrichment analysis |
如图5中肝脏组织苏木精伊红染色显示, 夜晚光照组肝细胞明显肿大, 胞浆稀疏呈网状, 半透明, 出现肝细胞水肿。 与夜晚暴露1 h相比, 暴露3 h肝细胞水肿更明显。 表明夜晚光照可引起明显的肝细胞水肿, 且长时间夜晚光照危害更大, 提示肝脏出现炎症反应。 如图6所示, 肝脏组织免疫荧光染色显示, 夜晚光照引起促炎因子白介素17A和粒细胞-巨噬细胞集落刺激因子表达上调。 荧光定量分析结果显示, 夜晚3 h光照组较1 h光照组促炎因子表达显著升高; 同时与照射蓝光相比, 照射白光时促炎因子的上调更显著。 以上结果合并表明夜晚光照可引起免疫细胞浸润和炎症因子表达上调, 且夜晚白光及长时间光照危害更大。
 | 图5 肝脏组织苏木精伊红染色(n=6) 注: Ctl: 对照组; LAN-WL-1h: 夜晚白光1 h组; LAN-WL-3h: 夜晚白光3 h组; LAN-BL-1h: 夜晚蓝光1 h组; LAN-BL-3h: 夜晚蓝光3 h组Fig.5 Hematoxylin-eosin staining of liver tissue Note: Ctl: Control group; LAN-WL-1h: White light exposure at night for 1 hour group; LAN-WL-3h: White light exposure at night for 3 hours group; LAN-BL-1h: Blue light exposure at night for 1 hour group; LAN-BL-3h: Blue light exposure at night for 3 hours group |
 | 图6 肝脏组织免疫荧光染色 (a): 代表性免疫荧光图; 绿色标记白介素17A蛋白, 红色标记GM-CSF蛋白, 蓝色标记细胞核; (b): 荧光信号定量分析(n=6; * * p< 0.01, * * * p< 0.001 vs. 对照组) 注: Ctl: 对照组; LAN-WL-1h: 夜晚白光1 h组; LAN-WL-3h, 夜晚白光3 h组; LAN-BL-1h: 夜晚蓝光1 h组; LAN-BL-3h: 夜晚蓝光3 h组; IL-17A: 白介素17A; GM-CSF: 粒细胞-巨噬细胞集落刺激因子; DAPI: 4’ , 6-二脒基-2-苯基吲哚Fig.6 Immunofluorescence staining of liver tissue (a): Representative immunofluorescence images; Green color represents interleukin 17 A(IL-17A), red color represents granulocyte-macrophage colony-stimulating factor (GM-CSF), and blue color represents 4’ , 6-diamidino-2-phenylindole(DAPI); (b): Quantitative analysis of fluorescence signals Note: Ctl: Control group; LAN-WL-1h: White light exposure at night for 1 hour group; LAN-WL-3h: White light exposure at night for 3 hours group; LAN-BL-1h: Blue light exposure at night for 1 hour group; LAN-BL-3h: Blue light exposure at night for 3 hours group; IL-17A: Interleukin 17A; GM-CSF: Granulocyte-macrophage colony-stimulating factor; DAPI: 4’ , 6-Diamidino-2-phenylindole |
生物钟系统与光环境同步对维持正常生理功能至关重要。 本研究采用夜晚光照模式, 正常饮食, 发现长期持续夜晚蓝光暴露导致肝脏生物钟紊乱。 与夜间活动的啮齿类动物相比, 采用白天活动的啮齿类动物沙鼠, 更接近于人类日常生活环境, 从而为转化研究提供了更好的模型。
昼夜节律系统对不同光谱的敏感性不同。 光在调节大多数动物的行为和生理方面起着核心作用, 有研究发现视锥细胞、 视杆细胞及自主感光视网膜神经节细胞三种细胞均参与昼夜节律调控, 其中视网膜神经节细胞接收来自视锥细胞和视杆细胞的突触输入, 产生成像视觉; 而自主感光视网膜神经节细胞表达一种特有的视色素— — 视黑蛋白, 视黑蛋白对约480 nm的蓝光最敏感, 使自主感光视网膜神经节细胞能够在没有视锥和视杆细胞输入的情况下对光作出响应, 产生非成像视觉, 两者共同参与调控代谢。 本研究发现夜晚照射白光或蓝光均会破坏昼夜节律, 引发肝脏炎症反应。 Nagai等[16]发现夜间照射蓝光能干扰小鼠视交叉上核的生物钟基因节律, 滤除蓝光后光照改变小鼠行为节律相位的能力下降一半, 且抑制蓝光能改善代谢异常。 证明光对代谢的调控由成像视觉通路和非成像视觉通路共同介导。
肝脏作为最具有节律的组织器官, 在代谢的调节中起到重要作用。 本研究通过分析夜晚光照对肝脏生物钟基因表达节律的影响, 发现夜晚光照引起肝脏组织生物钟基因Arntl、 Clock、 Per2、 Cry1、 Nr1d1、 Rorα 表达节律的不同程度破坏, 同时与暴露1 h相比, 暴露3 h对生物钟基因表达节律的影响更显著, 表现为对基因节律的几乎完全逆转。 Borck等[17]发现暴露在夜晚光照下的C3H/HePas小鼠出现肥胖和肝脏脂肪变性, 与Rev-erbα 的表达减少有关; 另外Rev-erbα 靶向miRNA的表达增加。 Fonken等[18]发现夜间暴露在昏暗光线(5 lux)下的小鼠Rev-erb在肝脏和脂肪组织中的表达降低。 本研究结果也发现夜晚长时间蓝光照射下Rev-erbα 的表达降低。 大多数生物体已经发展出内源性驱动的昼夜节律与日常明暗周期同步。 近年来, 人类已经从自然发生的太阳光循环转向人工照明。 这些结果意义重大, 证明环境光照的轻微变化可以改变昼夜节律和代谢功能。
每一种生物功能都需要能量管理, 因此新陈代谢是生命的重要组成部分。 第一批单细胞生物出现以来, 就需要保护免受环境损害, 从而导致免疫系统的进化。 因此新陈代谢和免疫从生命开始就交织在一起。 代谢系统和免疫系统是生存的最基本需求, 在整个物种进化中都是保守且高度整合的, 两者功能相互依赖[19]。 在高等生物中控制关键代谢和免疫功能单位是从共同的祖先结构进化而来的— — 脂肪体, 它包含了哺乳动物的肝脏、 造血系统和免疫系统的同源物[20]。 研究发现持续暴露于病原体或病原体相关成分可以破坏从果蝇到人类的全身代谢功能[21]。 大多数现代人目前的营养习惯和生活方式严重倾向于代谢超载和体力活动的减少。 早在2007年国际癌症研究机构就指出轮班工作可能致癌[22]。 还有研究发现生物钟基因敲除小鼠肺肿瘤组织的原癌基因c-Myc表达升高、 增殖能力增强, 出现代谢异常, 促进小鼠肺癌的发生发展[23]。 一些大型临床研究探索了夜班工作和代谢性疾病之间的关系。 一项纳入14万余例的前瞻性研究显示, 女性护士夜班工作每增加5年, 患2型糖尿病的风险增加31%[24]。 一项多中心研究发现熬夜或夜班工作导致昼夜节律紊乱从而加重急性心肌梗死患者急诊再灌注术后心肌损伤, 并导致长期临床预后不良。 在5年的平均随访期间, 与不上夜班的人相比, 经常上夜班与主要心血管不良事件的风险增加92%有关[25]。 本研究也发现夜晚光照主要影响免疫系统和内分泌系统, 可能引发感染性疾病、 癌症、 免疫疾病和心血管疾病。
脂肪组织和肝脏都有一个类似的结构组织, 包括代谢细胞(脂肪细胞或肝细胞)与免疫细胞(库普弗细胞或巨噬细胞), 都可以直接进入一个巨大的血管网络。 这两个组织形成了一个适合的环境, 在免疫和代谢反应之间进行持续和动态的相互作用[20]。 十多年前的一项研究发现, 肿瘤坏死因子-α 在肥胖小鼠的脂肪组织中过度表达, 首次明确了肥胖和慢性炎症之间的联系[26]。 肿瘤坏死因子-α 是一种促炎细胞因子, 可激活各种信号转导级联反应。 肝脏是一个独特的免疫器官, 具有独特的免疫微环境, 是全身固有免疫应答反应的重要场所[27]。 肝脏作为身体最大的免疫器官, 是巨噬细胞重要的储库, 肝脏中储存的巨噬细胞占了体内巨噬细胞的80%~90%。 还有一种“ 库普弗细胞” , 既是肝脏的卫士, 也是全身的保护神, 可吞噬、 消化对机体有害的物质, 或者交给其他免疫细胞进一步清除。 本研究中KEGG功能富集分析显示, 差异表达的基因主要集中在核因子κ B信号通路, 吞噬, 造血干细胞分化, 天然免疫信号通路和获得性免疫信号通路。 肝脏组织H& E染色也显示, 夜晚光照引起明显的肝细胞水肿, 提示肝脏有炎症反应。 肝脏组织免疫荧光染色显示, 夜晚光照引起促炎因子白介素17A和粒细胞-巨噬细胞集落刺激因子表达上调。 炎症和代谢信号之间的联系是一种微妙的平衡, 虽然有短期补偿和适应性措施来保持这种微妙的平衡, 而从长远来看, 结果往往是有害的[20]。
本研究存在一定的局限性, 与代谢和免疫相关的脂肪组织有待进一步研究。 尽管存在局限性, 本研究的发现意味着与夜晚长期光照相关的不良影响可以通过调控生物钟, 抑制炎症来缓解。 未来需要更多的研究来揭示夜晚光照对代谢和免疫的作用机制。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|