
{kind=link}
{kind=link}
{kind=link}
{kind=link}
青藏高原不同植被亚型的光谱特征分析
[牛洁琼1  , 任鸿瑞
, 任鸿瑞1 , 周广胜2, * ]
, 任鸿瑞]
|
|


作者简介: 牛洁琼,女, 2000年生,太原理工大学测绘科学与技术系博士研究生 e-mail: jieqiongniu@163.com
光谱曲线是植被生理性状的重要表征, 研究植被光谱对监测植被的生长状况和健康状态具有重要意义。 当前植被光谱研究多对同一植被型内不同植被进行分析, 鲜有系统地整合不同植被亚型光谱特征的报道。 近年来青藏高原的生态格局发生了重大变化, 为实现基于青藏高原大范围尺度的植被光谱特征研究工作, 探讨不同植被亚型的光谱可分离性, 基于高分辨率Sentinel-2A/B遥感影像, 通过一阶微分光谱和植被指数对青藏高原19种植被亚型的光谱曲线进行分析。 结果表明: (1)植被亚型的光谱曲线在近红外波段表现出明显差异, 其反射率值可用于区分不同植被亚型; (2)一阶微分处理能够明确植被三边特征, 植被的一阶微分光谱具有规律性, 在红边处差异显著; (3)不同植被亚型的植被指数与红边参数变化相一致, 植被指数虽然数值偏小但具有区分性; (4)综合运用光谱反射率值、 三边参数和植被指数, 可区分不同植被亚型。 在植被亚型的尺度上分析了不同植被的光谱曲线特征, 实验结果可为植被亚型光谱数据库的完善提供数据支撑, 对青藏高原植被亚型精细识别工作具有实际意义。
The spectral curve is an important representation of vegetation physiological traits. The study of vegetation spectra is significant for monitoring vegetation growth and health status. Current research on vegetation spectra mainly analyzes different vegetation within the same vegetation type, and there are few reports on the systematic integration of spectral characteristics of different vegetation subformations. The ecological pattern of the Tibetan Plateau has changed significantly in recent years. To conduct research on vegetation spectral characteristics on a large scale in the Qinghai-Tibet Plateau region and to explore the spectral separability among different vegetation subformations based on high-resolution Sentinel-2A/B remote sensing images, the spectral curves of 19 vegetation subformations were analyzed by using the first derivative of the spectrum and vegetation indices. The results showed that: (1) The spectral curves of vegetation sub-formations exhibit significant differences in the near-infrared band, and their reflectance value can be used to distinguish different vegetation sub-formations; (2) First-order derivative processing can elucidate the three-edge features characteristics of vegetation, the first-order derivative spectra of vegetation are of certain regularity, with significant differences in the red edge; (3) Vegetation indices of different vegetation subformations are consistent with changes of the red edge characteristics parameter, and the vegetation indices are discriminative, although their values are relatively small;(4) By combining near-infrared peaks, three-edge characteristics parameters, and vegetation indices, different vegetation subformations can be distinguished. This study analyzed the spectral curve characteristics of different vegetation at the vegetation subformations scale. The experimental results can provide data support for improving the vegetation subformations spectral database and have practical significance for the fine identification of vegetation subtypes on the Qinghai-Tibet Plateau.
植被是覆盖地表的植物群落的总称, 它既是陆地生态系统的主体, 也是人类重要的环境资源和物质资源, 对碳平衡调节、 水文循环、 土壤保持和维持气候稳定等至关重要[1, 2]。 植被亚型是植被分类中的辅助级单位, 反映了植被的生态特征及其生长的区域气候和环境条件, 能够为研究一定地理区域和植物区系地区内的植物群落提供参考[3]。 准确描绘植物群落特征、 探讨不同类型之间的差异对植被资源的合理保护利用和人类社会的可持续发展具有重要意义。
区别于土壤、 水体等其他地物, 植被的光谱反射特征独特, 由其化学特征和形态学特征共同决定, 与其自身的生理性状表现有着密切的关系。 受叶片色素及细胞构造不同的影响, 植被的光谱曲线在不同波段有明显不同的特征, 可表征生长状况的众多信息, 如覆盖度和生物量等。 对植被光谱进行研究, 可以分析植被的健康状况和生长条件, 对监测植被群落动态、 了解植被对环境的响应、 开展植被分类识别工作等研究有着重要作用[4, 5]。 近年来, 遥感技术已被广泛应用于植被光谱特征研究工作, 国内外许多学者对植被的光谱特性[6, 7, 8]、 不同影响因素下的光谱特征[9, 10, 11]、 植被光谱识别和分类[12, 13, 14]、 植被光谱库的建立[15, 16]等开展了一系列的研究, 细致地分析了植被光谱特征规律, 为植被变化的遥感监测提供了大量基础数据。
青藏高原地区环境之独特可与地球南、 北极并列, 被称为“ 第三极” , 是我国重要的生态安全屏障[17]。 随着全球气候持续变暖, 青藏高原的生态环境和水循环格局发生了重大变化, 高原冰川退缩、 积雪融化、 草原冻融等问题频出, 进一步凸显了高原生态系统的脆弱性, 因此, 亟需对青藏高原区域的环境变化进行系统研究并总结出相关科学对策[18]。 植被是气候变化的敏感指示器[19], 研究青藏高原的植被分布特征能够明确区域内植被活动的变化以及高寒生态系统对气候系统的反馈。 光谱曲线作为植被生理性状的表征, 能够揭示植被的覆盖特征和群落结构等信息, 开展植被光谱特征研究工作是研究植被分布的重要组成部分。 对青藏高原区域的典型植被进行光谱特征提取与分析, 尝试揭示不同植被光谱曲线的规律, 可为青藏高原区域典型植被的识别分类提供科学依据, 对青藏高原植被多样性监测和生态环境保护规划意义重大。 目前已有研究对青藏高原内森林[20]、 灌丛[21]和草地[22]等植被类型的光谱特征进行了测定和分析, 但以往研究多基于小区域开展, 研究区域常为青藏高原内局部地区, 基于青藏高原大范围尺度的植被光谱研究较少, 同时, 以往研究中研究样地设置一般偏少, 数据缺乏普适性, 难以应用于大区域尺度的植被光谱特征研究工作。 此外, 已有研究多以同一植被型为研究对象, 如仅对高寒灌丛、 高寒草甸等植被型内的不同植被进行光谱特征分析, 系统地整合不同植被类型的光谱曲线规律的工作仍然较少, 尤其是细分到植被亚型尺度上的研究。 青藏高原内植被类型多样, 不同植被亚型的植被种类和群落结构表现出较大差异, 有必要获取相应的光谱特征来完善不同植被亚型的特征光谱库, 为基于植被亚型尺度进行青藏高原大范围植被遥感监测提供数据支撑。
本研究以青藏高原19类植被亚型为研究对象, 基于谷歌地球引擎(google earth engine, GEE)平台获取高分辨率Sentinel-2A/B遥感影像数据, 根据波段信息提取不同植被在生长旺盛期的光谱曲线, 构建了包括归一化植被指数(normalized difference vegetation index, NDVI)在内的12个常用植被指数, 同时对植被原始光谱曲线和经过一阶微分处理后的光谱曲线进行分析, 探讨了不同植被亚型的光谱可分离性, 旨在揭示不同种类之间的光谱特性差异和分类规律, 以期为植被亚型尺度上的青藏高原植被光谱识别和植被光谱库的完善提供依据。
青藏高原(25° 59'30″— 40° 1'0″N, 67° 40'37″— 104° 40'57″E)位于亚洲中部, 平均海拔约4 400 m, 总面积约308.34万km2, 在我国境内的面积约为258.09万km2 [23]。 青藏高原地势呈现西高东低的特点, 由东南向西北, 随着地势升高, 高原各地年均温由东南部的20 ℃以上递降至西北部的-6 ℃以下, 年降水量也相应地由2 000 mm以上渐减至50 mm以下。 青藏高原植被类型多种多样, 包括森林、 草原、 草甸、 高山植被、 灌丛等, 草原、 草甸和高山植被面积较大, 约占青藏高原区域的66%以上。 空间分布上, 藏南部和东南部地区多分布着高大的乔木, 灌木类植被也主要分布在青藏高原东南部。 中部地区植被类型以高寒草原为主, 中东部以高寒草甸为主, 藏西北主要分布着荒漠植被[24]。
1.2.1 遥感数据
遥感数据使用欧洲航天局设计开发的Sentinel-2系列卫星遥感影像。 Sentinel-2适用于海陆环境, 是高分辨率多光谱成像卫星, 搭载多光谱成像仪(MSI), 覆盖4个10 m、 6个20 m及3个60 m空间分辨率, 共有从可见光到短波红外的13个波段。 本研究使用GEE平台提供的地表反射率产品, 该2A级产品已经过了大气校正和正射校正, 只需利用Cloud Probability数据集进行影像去云操作。 基于GEE平台获取2020年7月15日至2020年8月15日, 完全覆盖青藏高原区域的Sentinel-2A/2B影像。
| 表1 Sentinel-2波段信息 Table 1 Sentinel-2 band information |
1.2.2 样点数据
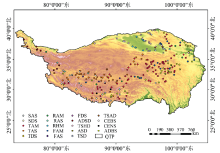
本研究使用已公开的青藏高原植物群落样方数据集作为样点数据[25], 该数据集共包含338个样地信息和758个样方记录, 提供了青藏高原温性草原和荒漠植被区域以及高寒植被区域的资料。 综合考虑采样点分布和植被类型, 选取279个样地数据进行分析, 样地分布在青藏高原三江源地区、 藏南山地、 阿里地区、 柴达木盆地, 以及祁连山、 阿尔金山、 昆仑山等高山区域, 植被类型以高寒草甸、 草原、 灌丛和荒漠以及温性草甸、 草原和荒漠为主。
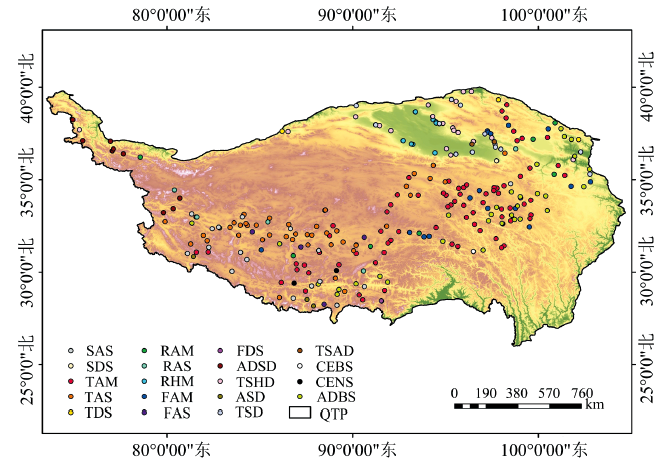 | 图1 研究样方数据集空间分布Fig.1 Distribution of the sample sites on the Qinghai-Tibet Plateau |
利用已有样地的坐标定位信息, 基于处理后的Sentinel-2A/B影像提取样本点对应像元的波段反射率值, 计算得到植被的平均光谱反射率, 以绘制光谱曲线。
1.3.1 一阶微分处理
光谱微分技术是最常用的光谱增强技术之一, 它可以提高信噪比, 有效去除背景干扰, 消除由于大气效应引起的噪声现象, 增强绿色植物的光谱特征差异。 本研究对获取的植被光谱曲线进行一阶导数变换并分析其红边、 黄边、 蓝边特征。 光谱反射曲线的一阶导数曲线公式为[26]
式(1)中, λ i为波段i的波长值; ρ (λ i )为波长λ i的光谱反射率值; ρ '(λ i)为波段i的一阶微分值。 为定量反映不同植被类型间的光谱特征差异, 提取9个常用的一阶导数光谱特征参数[27], 包括蓝边斜率、 蓝边位置、 蓝边面积以及相应的黄边和红边参数。
1.3.2 植被指数提取
植被指数是定量和定性评价植被覆盖度和生长状况等的有效变量。 本研究基于Sentinel-2A/B遥感影像数据, 根据波段信息构建了包括NDVI在内的12个常用的植被指数作为辅助分析。
| 表3 一阶导数光谱特征参数 Table 3 Characteristic parameters of the first derivative spectrum |
| 表4 本研究使用的植被指数 Table 4 The vegetation indices used in the study |
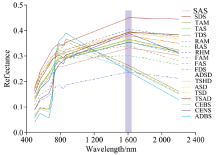
利用Sentinel-2A/B遥感影像和波段反射率值数据得到研究区内不同植被类型的光谱曲线如图2所示。
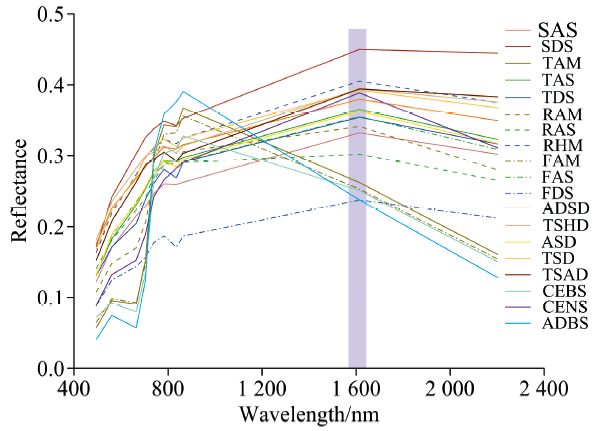 | 图2 各植被亚型的原始光谱曲线Fig.2 The original spectral curves of 19 vegetation subformations |
受地形和气候条件的影响, 青藏高原区域内的植被普遍生长矮小, 叶片发育较少, 植被光谱特征不明显, 但仍具有大多数健康绿色植被光谱的“ 峰和谷” 的特征。 在380~780 nm波段的可见光范围内, 不同植被类型的光谱曲线在波长550 nm附近均表现出明显的反射峰, 即“ 绿峰” 现象; 在波长675 nm附近光谱曲线吸收谷“ 红谷” 比较明显; 随后在680~750 nm的波长范围内, 光谱反射率随波长的增加而急剧升高, 光谱曲线较为陡峭, 形成植被光谱特有的“ 红边” 特征。 近红外波段范围内是一个相对平坦的区域, 光谱反射率一直保持较高水平, 在780~830 nm波长内光谱反射率缓慢降低, 随后有所增加并在波长860 nm附近出现一个小的反射峰。 波长1 500 nm后, 除TAM、 FAM、 CEBS和ADBS在波长1600nm附近光谱反射率较低, 光谱曲线总体下降, 其余植被类型的光谱曲线均呈现出先增大后减小的趋势, 在1 600 nm处出现一个反射波峰后逐渐减小。
19类植被光谱曲线的总体走势相近, 但区分程度各不相同。 在可见光波段内, 各植被类型的反射率差异并不明显。 在近红外区, 植被反射率差异较大, 波长1 600 nm处各植被的光谱反射率差异明显, 平均光谱反射率由高到低依次为SDS> RHM> TSHD> TSAD> TSD> CENS> ADSD> TAS> ASD> FAS> TDS> RAM> SAS> RAS> TAM> FAM> CEBS> ADBS> FDS, 其中SDS的光谱反射率最大, 达到了44.99%, 而最小者FDS仅为23.76%。 少数植被的反射率值较为接近, 如TSAD(39.42%)和TSD(39.22%), TAS(36.52%)和ASD(36.28%), 以及FAS(35.52%)和TDS(35.42%), 但TSAD在可见光波段内的光谱反射率一直低于TSD, ASD在830 nm附近的光谱反射率明显低于TAS, 此外, 根据750~900 nm波长范围内反射率的不同也能辨识FAS和TDS。 总体而言, 各植被亚型的光谱曲线在近红外区域明显不同, 结合不同波长范围内的光谱曲线差异, 可以对不同植被亚型进行区别与判识。
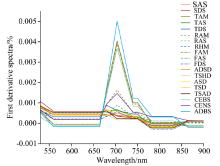
为使植被的光谱曲线变化特征更为突出, 对19种不同植被的原始光谱曲线进行了一阶微分处理, 截取了光谱曲线差异显著的区域(490~900 nm)用于分析(图3), 同时根据一阶微分光谱曲线提取9个常用的一阶导数光谱特征参数。
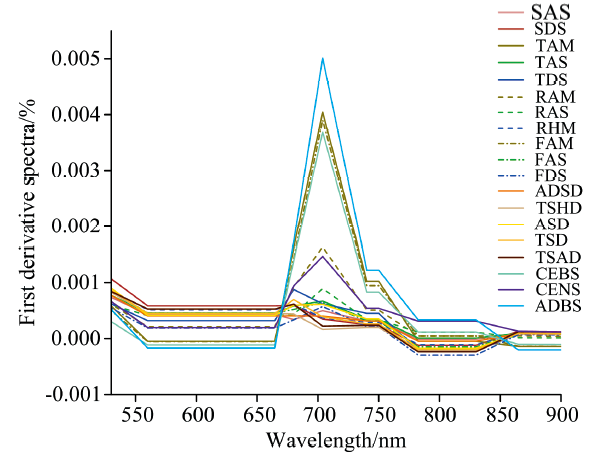 | 图3 各植被亚型的一阶微分光谱曲线(490~900 nm)Fig.3 The first derivative reflectance spectra of 19 vegetation subformations (490~900 nm) |
不同植被类型的一阶微分光谱曲线总体趋势基本相似, 存在较为明显的植被特有的吸收和反射特征。 在550 nm附近各植被类型的一阶微分光谱曲线均出现极值, 呈现反射峰特征。 在490~750 nm波长范围内, 植被的一阶微分值均为正数, 表明植被的光谱反射率与波长呈正相关。 在680~750 nm范围内, 一阶微分曲线出现最大值, 即该波段范围内光谱反射率增幅达到最快, 与原始光谱在“ 红边” 区域反射率快速上升这一特征相一致。 780 nm后, 一阶微分光谱曲线变化趋于平缓。
对比各植被类型的一阶微分光谱特征参数, SDS的蓝边斜率(0.001 06)和蓝边面积(0.033 09)最大; ADBS蓝边参数最小, 仅为0.000 30和0.006 68。 同植被型中, 高寒草甸的蓝边参数明显小于其他植被亚型, 而高寒草原的蓝边参数较大。 荒漠类植被的蓝边参数较为接近, 分别位于0.000 8和0.025附近。 灌丛类植被中, CENS的蓝边参数远大于CEBS和ADBS, 数值约为其余两种亚型的两倍。
19种植被亚型的黄边参数中, ADBS的黄边斜率和SDS的黄边面积最大, 数值分别为0.001 27和0.046 87; FDS黄边参数最小, 仅有0.000 30和0.019 43。 不同植被型中高寒草甸的黄边参数明显大于同植被型中其他亚型。 荒漠类植被的黄边参数呈递增趋势。 灌丛类植被的黄边参数中, CENS的黄边斜率远小于其他两类。
“ 红边” 是植被光谱曲线研究最重要的部分, ADBS的红边斜率和红边面积最大, 分别达到了0.005 0和0.220 0; 而TSHD的红边斜率仅有0.000 3, 红边面积0.013 9, 为植被亚型中的最小值。 不同于蓝边参数, 丛生草类、 根茎草类和杂类草植被型中, 高寒草甸的红边参数为同类别中最大, 高出同类别植被亚型2~3倍。 荒漠类植被的红边参数均较低, 红边斜率处于0.000 3~0.000 6范围内, 红边面积在0.01~0.03之间。 灌丛类植被的红边参数较高, CENS的红边参数远低于CEBS和ADBS, 但仍明显高于其他的植被亚型。
图4显示, 19种植被光谱的红边位置平均值约为711 nm, SAS、 TAM、 TAS、 RAM、 RAS、 FAM、 FAS、 FDS、 ADSD、 ASD、 CENS、 CEBS和ADBS的红边位置较711 nm波长有不同程度的增加, 其中ADBS“ 红移” 幅度最大; 而SDS、 TDS、 RHM、 TSHD、 TSD和TSAD存在“ 蓝移” 现象, TSD“ 蓝移” 幅度最大。 本研究中ADBS植被亚型多为金露梅灌丛, 相比其他植被, 金露梅灌丛较茂盛, 在生长旺盛期其叶片叶绿素含量较高, 对红光的吸收强, 红边位置向长波方向移动, 因此“ 红移” 幅度更大。 存在“ 蓝移” 现象的植被亚型多为荒漠类植被, 荒漠类植被中叶绿素含量相对更少, 导致红光反射增强, 植被存在一定的红边“ 蓝移” , 红边位置都靠近短波方向; RHM作为盐生植被, 分布在盐碱化土壤上, 高浓度盐胁迫对植被根系生长表现出明显抑制作用, 红边位置易出现较大幅度的“ 蓝移” 现象。
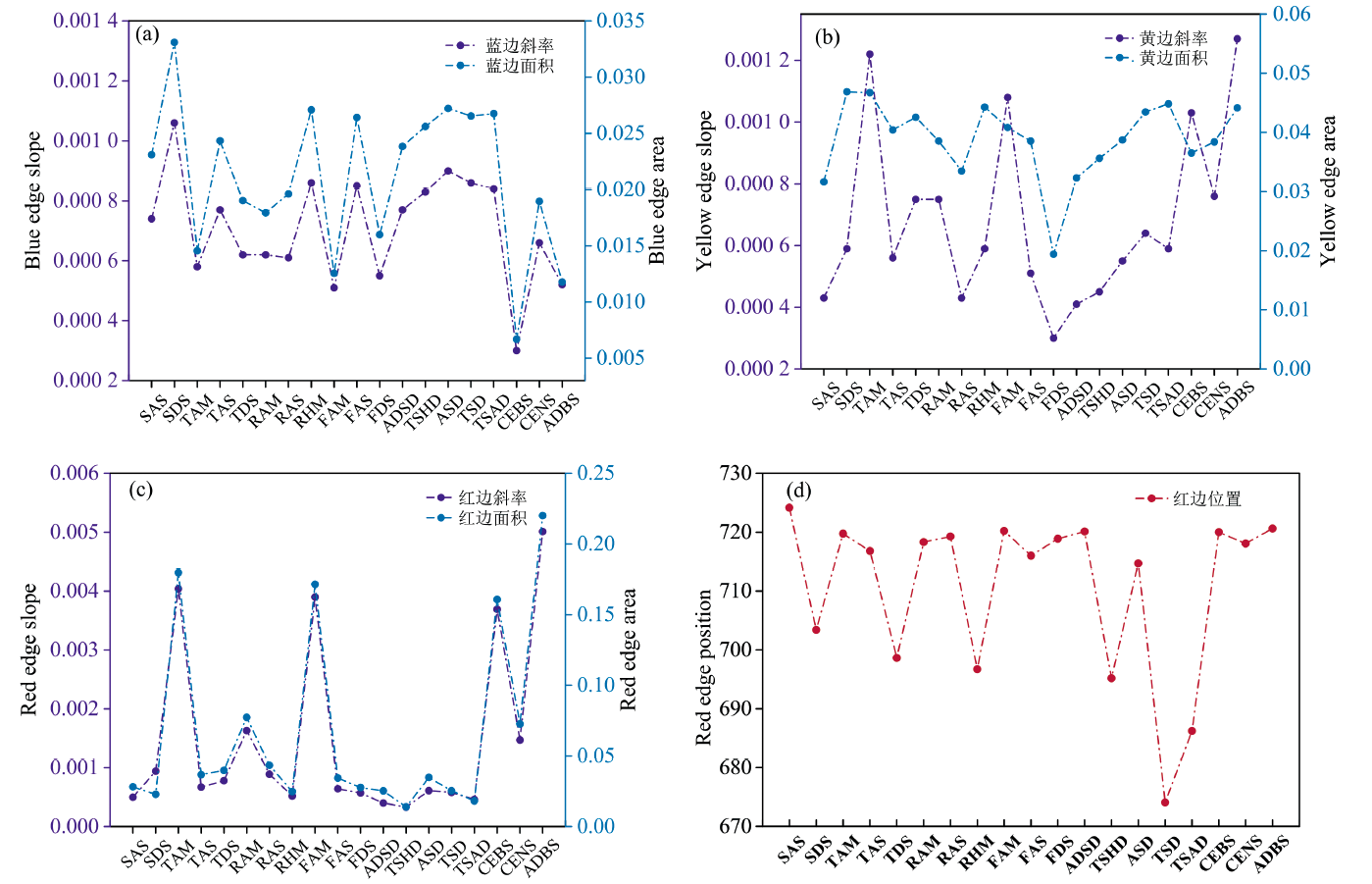 | 图4 各植被亚型的蓝边参数(a)、 黄边参数(b)、 红边参数(c)及红边位置(d)Fig.4 Parameters of three sides [blue (a), yellow (b) and red (c)] and red edge (d) positions of 19 vegetation subformations |
总体而言, 各植被类型在蓝边、 黄边和红边的光谱特征变化各不相同, 在红边处的差异最为明显, 可区分性较强。
本研究选取并计算了12个常用的植被指数, 植被指数的具体数值见表5。
| 表5 不同植被亚型的植被指数 Table 5 The vegetation indices of 19 vegetation subformations |
青藏高原地区寒冷干旱, 植被生长较为矮小, 覆盖度也较低, 植被指数数值总体偏小。 以归一化植被指数(NDVI)为例, 除红边参数较大的四类植被亚型的NDVI值大于0.5外, 其余植被亚型的NDVI值均位于0.1附近, 小于正常情况下覆盖度较大的绿色植被。
植被指数与红边参数具有一致的变化特征。 在19种植被亚型中, ADBS的植被指数数值均为最大, 这与其红边参数最大这一特征相符合。 ADBS群落茂盛, 覆盖度高, 叶片叶绿素含量高, 长势较好, 其对应的表征植被生长状况的植被指数数值较大。 ADSD的红边归一化植被指数(RNDVI)和红边比值植被指数2(RRI2)以及SDS的三角植被指数(TVI)值是最小的, 其余的植被指数最小值出现在TSHD类别中。 ADSD和TSHD是高寒荒漠植被, 荒漠植被生长环境极端寒冷严酷, 植被低矮稀疏, 多贴地生长, 其叶面积明显缩小, 直观反映为荒漠类植被的各植被指数明显小于其余植被亚型。
灌丛类植被的植被指数数值较高, CEBS的各植被指数稍低于其他两类灌丛, 这可能是由于本研究中CEBS为刺柏灌丛, 叶面积小于包括金露梅灌丛和杜鹃花灌丛在内的其他两类阔叶灌丛。 在丛生草类、 根茎草类、 杂类草植被型中, 高寒草甸的植被指数数值为同类别中最大, 高寒草甸属于高寒半湿润半干旱气候, 土壤水分含量相对高, 利于植被生长, 高寒草原气候严寒干旱, 植被覆盖度相比高寒草甸小, 受生境条件和植被盖度的影响, 高寒草甸的植被指数数值较高。 盐生草甸分布地区的土壤多为盐碱土和盐化草甸土, 盐碱浓度较高的环境会影响植物根系的吸水能力和叶绿素合成, 导致植被红边位置“ 蓝移” , 表征着植被叶绿素含量减少, 因此RHM对应的各植被指数值在同植被型中为最小。
植被指数能够评价植被覆盖度、 反映植被生长状况, 是植被遥感监测的基础, 本研究中19类植被亚型的植被指数虽然数值偏小, 但仍具有一定的区分性, 可用于植被光谱识别中的辅助分析。
植被亚型的光谱曲线具备植被特有的光谱反射特征, 在可见光波段, 受植物叶片中的叶色素的影响, 绝大多数植物在450和670 nm波长附近存在波谷。 在490~600 nm波长范围内, 不同的植被均出现了较明显的反射峰, 这一波长范围是植物叶片光合作用利用较少的区域, 辐射反射率较强。 在680~750 nm波长范围内, 植被的光谱反射率快速升高, 呈现出“ 红边” 区域高反射的特征。 在近红外波段, 不同植被亚型的反射率差异较为明显, 可利用波长1 600 nm处光谱反射率的大小顺序对植被亚型进行区分, 对于少数反射率值较为接近的植被亚型, 也可通过其在其他波长范围内的大小顺序进行区分。
一阶微分处理应用于多光谱数据已有先例, 如用于光谱识别、 地物分类和目标检测等多种任务。 对多光谱数据进行一阶微分计算, 可以降低背景噪声并增强光谱数据特征, 能够更好地表达地物的真实情况, 突出细节信息[28]。 本研究采用一阶微分处理对植被光谱进行分析, 并通过三边参数反映植被亚型的光谱差异。 植被亚型的三边参数具有一定的规律性。 灌丛类植被的蓝边参数明显低于其他草本植物, 而红边参数则远大于其他植被亚型, 这可以作为灌丛类植被区别于草本植物的一个典型特征。 同灌丛类植被中, CENS的蓝边参数大于其他两类灌丛, 但红边参数最小, 可据此区分CENS。 同植被型中, 高寒草甸的红边参数显著高于高寒草原和荒漠草原, 这与已有研究[29]的结论相一致, 从高寒草甸到高寒草原到荒漠草原, 随着植被盖度等生物量信息减少, 红边参数呈现逐渐减小的趋势。 同时, 盐生草甸的红边参数也要小于其余植被亚型, 这符合已有的研究结论, 研究证实盐分影响是多数植物生长的抑制因子, 土壤盐渍程度的增加会导致植被红边位置“ 蓝移” 和红边参数的下降[9]。 TAM、 FAM、 CEBS和ADBS的植被群落较为茂盛, 生长状况较好, 红边参数远高于其他植被亚型, 在光谱曲线上表现为红边处这四类植被的曲线相对更为陡峭, 可分离性较强。
植被指数整体变化趋势与红边参数特征变化趋势一致, TAM、 FAM、 CEBS和ADBS的植被指数值较高, 其中, ADBS的植被指数值均为最大, 符合其红边参数最大这一特征; 作为红边参数最小的植被亚型, TSHD的9个植被指数值均为最小。 本研究中19类植被亚型的植被指数具有一定的区分性。 灌丛类植被的植被指数数值明显高于其他植被型, 而荒漠类植被的植被指数数值整体偏低。 同植被型中, 高寒草甸的植被指数数值明显高于高寒草原和荒漠草原。 在高寒草地中, 土壤水分和土壤温度的大小顺序依次为高寒草甸> 高寒草原> 荒漠草原, 已有研究证实植被指数NDVI在增湿的条件下呈增加趋势, 这一趋势以高寒草甸表现最为明显[30], 这与本文高寒草甸的植被指数数值高于其他两类草地的特征一致。
本研究通过植被原始光谱曲线和一阶微分光谱曲线进行不同植被亚型的光谱特征分析, 探讨了青藏高原大范围尺度上不同植被亚型的光谱可分性, 研究表明青藏高原区域不同植被亚型光谱具有较强的可分离性, 为青藏高原植被光谱库的丰富提供了数据支撑, 为在青藏高原大范围区域进行植被亚型尺度的光谱识别工作提供了实际依据。 但本研究仍可能存在一些局限性: (1)本研究为保证采样点与影像数据的时空一致性, 获取了2020年7月— 8月的Sentinel-2影像, 植被在不同季节的光谱特征变化仍有待深入研究。 (2)本研究对光谱特征进行分析采用了最常用的光谱增强技术, 即一阶微分处理技术, 该方法能够很好地消除背景噪声的影响。 但其他方法也可用于对原始光谱曲线进行处理, 如连续统去除处理等。 未来在对植被的原始光谱进行处理时, 可以考虑使用更多的光谱处理技术, 为光谱曲线特征数值的比较提供更多参考。 此外, 可以在研究中加入样地实测光谱数据用于分析, 以获取更多的植被光谱反射率信息, 增强不同植被亚型之间的光谱区分度。 同时, 可以基于多时相的遥感影像对植被亚型的动态变化进行研究, 实现不同植被亚型、 同一植被亚型在不同时期的光谱特征差异的分析。
基于高分辨率Sentinel-2A/B遥感影像提取了植被的光谱特征, 根据波段特征构建了12个常用的植被指数, 通过对不同植被亚型的原始光谱、 一阶微分处理光谱进行差异分析, 得到如下结论:
(1)青藏高原区域的植被光谱具有“ 峰和谷” 的特征。 在近红外波段, 植被光谱曲线可区分性明显, 波长1600nm处各植被的平均光谱反射率由高到低为SDS> RHM> TSHD> TSAD> TSD> CENS> ADSD> TAS> ASD> FAS> TDS> RAM> SAS> RAS> TAM> FAM> CEBS> ADBS> FDS, SDS的反射率值最大, FDS的反射率值最小, 偶有反射率值较为接近的植被, 结合其他波长范围内的光谱曲线差异, 仍可进行有效区分。
(2)植被的一阶微分光谱曲线具有一定的规律性, 在红边处的差异最为明显。 同植被型中, 高寒草原的红边参数相对最小, 高寒草甸的红边参数为最大; 荒漠类植被的红边参数整体偏低; 灌丛类植被的红边参数处于较高水平, 其中ADBS的红边参数最大。
(3)19种植被亚型的植被指数数值偏小, 其变化与红边参数变化趋势一致。 TAM、 FAM、 ADBS和CEBS的植被指数数值明显高于其他植被亚型, 其中ADBS的12个植被指数值均为最大, 符合其红边参数最大这一特征。
(4)19种植被亚型的原始光谱曲线和一阶微分光谱曲线具有较强的可区分性, 利用不同植被亚型光谱反射率值的显著差异, 辅以三边参数和植被指数特征, 可以实现不同植被的有效分类和识别。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|