


{kind=link}
{kind=link}
{kind=link}
毛竹向杉木林扩张不同阶段叶面积指数地面高光谱遥感模型研究
[李聪慧1, 2  , 李宝银
, 李宝银1, * , 毛振伟3 , 李璐飞3 , 余坤勇4 , 刘健4 , 钟全林1 ]
, 李宝银, 毛振伟|
|


作者简介: 李聪慧, 女, 1987年生, 福建开放大学理工学院讲师 e-mail: conghui-li@foxmail.com
毛竹向杉木林扩张会引发“林退竹进”、 林权纠纷等生态与经济问题。 利用遥感有效反演毛竹演替过程对于科学管控森林资源意义重大。 为了揭示地面高光谱在毛竹扩张不同阶段叶面积指数(LAI)反演的有效性, 以毛竹在杉木林中扩张为研究对象, 沿毛竹扩张方向设置四类杉竹比例样方, 模拟扩张Ⅰ、 Ⅱ、 Ⅲ、 Ⅳ阶段, 根据原始光谱及开方、 对数、 倒数、 一阶微分、 二阶微分等10种光谱变换数据与不同扩张阶段LAI相关性选取特征波段, 构建归一化植被指数(NDVI)、 黄光波段指数(YI)等7个LAI显著相关植被指数, 分别建立光谱信息与植被指数的五类单因素回归模型, 基于植被指数利用神经网络、 决策森林回归、 贝叶斯线性回归和线性回归四种机器学习方法构建多因素回归模型, 探讨毛竹向杉木林不同扩张阶段LAI高光谱反演模型适用性。 结果表明: 原始光谱的一阶微分、 二阶微分、 对数一阶微分和倒数一阶微分四类微分变换能够丰富光谱信息, 更好表征毛竹向杉木林的扩张过程; 植被指数中, YI与毛竹LAI相关系数最大, 表现出毛竹扩张过程的高敏感性, NDVI反演效果最佳, 但基于传统植被指数建模的总体反演效果不佳; 基于微分变换光谱的二次多项式和幂指数回归模型在各扩张阶段表现更佳, 神经网络算法在LAI反演方面优于其他机器学习算法, 总体来看, 基于光谱变换的传统回归算法优于机器学习建模方法。 扩张第Ⅲ阶段基于对数倒数一阶微分变换光谱构建的模型( y=5.291 4e183.76 x)拟合效果最佳, 建模集和验证集 R2分别为0.735和0.742, RMSE分别为0.733和0.468, nRMSE分别为14.0%和9.9%, 建议毛竹扩张管控选在竹林各半的混交林中。 系统开展毛竹不同扩张阶段LAI高光谱反演模型的创新分析, 将为科学营林造林管理奠定基础。
The expansion of Phyllostachys edulis (Moso Bamboo) into Cunninghamia lanceolata (Chinese fir) forest has led to ecological and economic problems such as “forest retreat and bamboo advance” and forest rights disputes. The use of remote sensing to effectively invert the succession process of Moso bamboo is of great significance for scientific control of forest resources. To reveal the effectiveness of canopy hyperspectral inversion of leaf area index (LAI) at different expansion stages of Moso bamboo to Chinese fir forest, four types of sample squares in mixed forest, which were divided based on a percentage of Moso bamboo were set to simulate expansion stages Ⅰ, Ⅱ, Ⅲ and Ⅳ along the expansion direction. Meanwhile, to explore the applicability of LAI hyperspectral inversion models for different expansion stages of Moso bamboo to Chinese fir forest, five types of single-factor regression models were established based on the characteristic wavebands which were chosen by the correlation between the original spectra, 10 spectral transformations such as open square, logarithmic, inverse, first-order differential and second-order differential and LAI at different expansion stages, and seven LAI significantly related vegetation indices such as normalized difference vegetation index (NDVI), yellowness index (YI) and so on. Multi-factor regression models were established based on the vegetation indices using four machine learning methods: neural network, decision forest regression, Bayesian linear regression, and linear regression. The results showed that the differential transform, including first-order differentiation ( R'), second-order differentiation ( R″), logarithmic first-order differentiation [(lg R)'] and inverse first-order differentiation [(1/ R)'] of the original spectrum ( R), could enrich the spectral information to characterize the expansion process better. Among the vegetation indices, YI had the highest correlation coefficient with LAI, showing high sensitivity to the expansion process, and NDVI had the best inversion effect. However, the overall inversion based on traditional vegetation index modeling was in effective. Moreover, the quadratic polynomial and power exponential regression models based on the differential transform spectra performed better in each expansion stage, while the inversion effect based on the vegetation indices was poor, and the neural network algorithm outperformed other machine learning algorithms. Traditional regression algorithms based on spectral transformations performed better than machine learning modeling methods. The model ( y=5.291 4e183.76 x) based on log-inverse first-order differential transform spectra fitted best in expansion stage Ⅲ with R2 of 0.735 and 0.742, RMSE of 0.733 and 0.468, and nRMSE of 14.0% and 9.9% for the modeling and validation sets, respectively. We suggest that the Moso bamboo expansion control should be selected in mixed forests of half of each species. Innovative analysis of LAI hyperspectral inversion modeling of Moso bamboo at different expansion stages will provide a basis for scientific silvicultural management.
20世纪80年代以来, 毛竹(Phyllostachys edulis)以其生长速度快、 再生能力强、 笋材两用、 涵养水源等经济、 生态和文化特性备受林农青睐, 其固碳量大[1]、 生长周期短、 改善土壤环境[2]的优势符合“ 双碳” 目标, 也被作为当前的林业支柱产业大力发展。 毛竹和杉木(Cunninghamia lanceolata)因立地条件相近, 在我国南方集体林区常相伴而生[3], 但毛竹快速出笋成竹的空间侵占、 鞭根养分主动掠夺及化学“ 他感” 抑制原生林生长的优生策略使杉木林出现“ 林退竹进” 现象[4], 亦对生物多样性和森林生态结构产生负面影响[5]。 尤其近年来新型合成替代材料行业发展及人力成本提升造成的竹林弃管, 更是日益加剧了林地边界增扩引发的林权负面纠纷, 严重影响南方集体林区森林资源经营管理稳定, 制约林权制度改革的完善与深化。
作为链接生物小循环过程中能量流动与物质循环的重要参考指标, 叶面积指数(leaf area index, LAI)在各类陆地植被模型构建中被广泛使用, 常用于描述植被冠层结构[6]。 利用遥感反演LAI监测植被生长过程是科技赋能农林业发展的高效手段, 地面高光谱丰富的波段信息为LAI反演提供了更为精细的数据支撑, 已为业界广泛使用[7, 8]。 毛竹扩张的过程会通过冠层光谱曲线中的“ 绿峰” 和“ 红谷” 特征变化体现出来, 由此会引起归一化植被指数、 氮素反射指数、 黄光波段指数等参数变化[9, 10], 为毛竹扩张过程的冠层LAI反演提供了可能。 但目前利用地面高光谱数据反演作物不同生长时期LAI的研究较多, 尚缺乏毛竹扩张原生林过程研究, 且较多研究仅从光谱变换或植被指数单方面入手, LAI反演模型构建缺乏综合传统分析和机器学习方法对单因素和多因素进行全面对比分析。 因而, 综合原始光谱、 变换光谱信息及敏感植被指数, 构建单因素和多因素的线性及非线性回归分析, 并结合机器学习方法筛选出毛竹扩张不同阶段LAI反演的最适模型成为研究重点, 也将进一步为毛竹向原生林扩张过程监测及森林资源科学经营提供数据支撑。
研究区选取福建省三明市永安天宝岩国家级自然保护区内毛竹扩张杉木林显著林区(图1), 地理边界为北纬25° 52'30″— 25° 52'39″, 东经117° 28'8″— 117° 28'29″, 海拔652~733 m; 中亚热带海洋性季风气候, 年均气温在23 ℃左右, 最低温度-11 ℃, 最高温度40 ℃; 年均降雨量2 000 mm左右, 湿度80%以上, 无霜期290 d左右。 研究区土壤以氧化土为主, 人工经济林为杉木、 毛竹高密度种植区并曾经过施肥、 采伐及林下植被清除等处理, 杉竹混交林分由40年林龄杉木及扩张近20年毛竹林形成, 少量伴生阔叶树种有木荷(Schima superba)、 南酸枣(Choerospondias axillaris)、 鹅掌楸(Liriodendron chinense)等。
 | 图1 研究区概况(中国地图-审图号GS(2019)1822号, 样地内数字为样方号)Fig.1 The location and sample setting of the study area (Map of China-Review No.GS(2019)1822 and the numbers in the sample plot represent the Plots No.) |
选取立地条件、 毛竹向杉木林扩张趋势相近样地4块, 沿毛竹向杉木林扩张方向设置10 m× 40 m调查样带15条, 各样带平均划分为4个10 m× 10 m面积大小样方(图1), 从毛竹向杉木扩张方向分别编号为1、 2、 3、 4, 根据毛竹扩张比例分别模拟扩张阶段Ⅰ 、 Ⅱ 、 Ⅲ 、 Ⅳ , 共计60个样方, 样地设置基本情况见表1。 随着扩张界面前移毛竹占比逐渐减少, 杉木占比逐渐增大。 根据各样方原始光谱信息及通过开方、 对数、 倒数、 一阶微分、 二阶微分等10种光谱变换数据, 由毛竹向杉木林扩张不同阶段LAI的相关系数确定敏感光谱波段, 用于构建敏感波段单因素回归模型及相关植被指数, 再以与不同毛竹扩张阶段LAI显著相关的植被指数分别构建单因素回归模型和多因素回归模型, 通过对比分析各类模型精度确定毛竹扩张杉木林不同阶段的LAI反演最优模型。
| 表1 样地设置基本信息表 Table 1 Basic information on site settings for different expansion stages of Phyllostachys edulis |
2021年7月, 选择试验区晴朗无云、 无风的天气, 利用产自仪思特光电技术有限公司(中国)的野外地物光谱辐射计(型号: ISI921VF-256, 光谱范围380~1 080 nm, 分辨率4 nm, 输出波段数256)进行毛竹冠层光谱数据采集。 每个样方随机选取3个测定点首先进行对照白板校正, 再各点收集10组冠层光谱数据, 保留390~995 nm波段以去除边缘波段影响, 并对30组数据取平均作为样方光谱反射率。 采用产自Li-COR公司(美国)的植物冠层分析仪(型号: LAI-2200)进行LAI数据测定, 每个样方测量3组重复数据取平均作为冠层LAI值。 应用Microsoft Office Home and Student 2019、 OriginPro 2021和IBM SPSS Statistics进行数据预处理、 相关性分析、 变换光谱处理及制图, Microsoft Machine Learning Studio进行机器学习建模, ArcGIS 10.4制作研究区位图。
植被指数对不同植被LAI的敏感性表现出高光谱反射率不同波段的差异性[11], 针对不同研究对象构建高敏感性植被指数是提升高光谱遥感反演精度的关键[12]。 根据前人关于LAI估测经验, 逐一验证与本研究LAI显著相关的经典植被指数, 最终选定归一化植被指数(normalized difference vegetation index, NDVI)、 绿色归一化植被指数(GNDVI)、 比值植被指数(RVI)、 氮素反射指数(NRI)[13]、 黄光波段指数(yellowness index, YI)[9]、 三波段修正归一化差分光谱指数(mNDSI)[14]、 新植被指数(NVI)[15]7个植被指数作为毛竹向杉木林扩张不同阶段LAI高光谱估测模型参数, 并以与毛竹向杉木林扩张LAI显著相关的558处波段(相关系数-0.359)作为“ 绿峰” 特征波段, 606处波段(相关系数-0.394)作为黄光特征波段、 676处波段(相关系数-0.416)作为“ 红谷” 特征波段以及800处波段(相关系数-0.247)作为近红外特征波段构建相关植被指数, 提高毛竹扩张不同阶段的LAI反演精度。
模型构建根据毛竹向杉木林扩张不同阶段的LAI实测值排序后, 等间隔取样, 选择2/3样本作为建模集, 其余1/3作为验证集检验模型精度。 采用与实测LAI显著相关的原始波段、 变换波段和植被指数建立单因素回归模型, 并利用相关的7个植被指数建立多因素回归模型。 以决定系数(coefficient of determination, R2)、 均方根误差(root mean square error, RMSE)、 归一化均方根误差(normalized root mean square error, nRMSE)评价模型精度[16], RMSE和nRMSE越小表明模型拟合效果越好。 单因素回归模型通过建立敏感波段与LAI之间的线性、 二次多项式、 指数、 幂指数和对数统计关系建模。 多因素回归模型通过决策森林回归(decision forest regression)、 贝叶斯线性回归(Bayesian linear regression)、 线性回归(linear regression)和神经网络(neural network)4种机器学习模型建模。
毛竹扩张会引起叶片、 冠层及立木环境的变化, 高光谱已被大量研究认为可以有效识别这些变化[17, 18, 19]。 在可见光范围内(图2), 受叶绿素影响毛竹光谱反射率出现510~560 nm之间的“ 绿峰” , 550 nm附近呈现反射率峰值, 同时, 受红光吸收作用出现680~750 nm之间的“ 红边” 效应, 650 nm附近表现为“ 红谷” 。 毛竹在可见光范围内的“ 绿峰” 和“ 红谷” 将有助于毛竹林的识别, 但在植物光谱曲线上普遍存在该现象[12], 在高光谱反射率上有效区分毛竹与原生林的关键在于物种间存在量级差异。 在“ 绿峰” 和“ 红谷” 之间, 毛竹向杉木林扩张各阶段存在明显差异, 且通过与前人关于杉木原始光谱的研究结果对比, 杉木“ 绿峰” 和“ 红谷” 光谱反射率均低于0.05, 而毛竹界于0.088~0.100之间, 相差较大利于有效区分, 可作为毛竹向杉木林扩张不同阶段的重点研究谱域。 此外, 由于叶肉细胞的“ 空腔” 结构使毛竹光谱反射率在大于760 nm的近红外波段明显高于可见光区, 在960 nm附近出现水分“ 吸收谷” , 毛竹向杉木林扩张的不同阶段在近红外区域呈现出更为明显的差异。
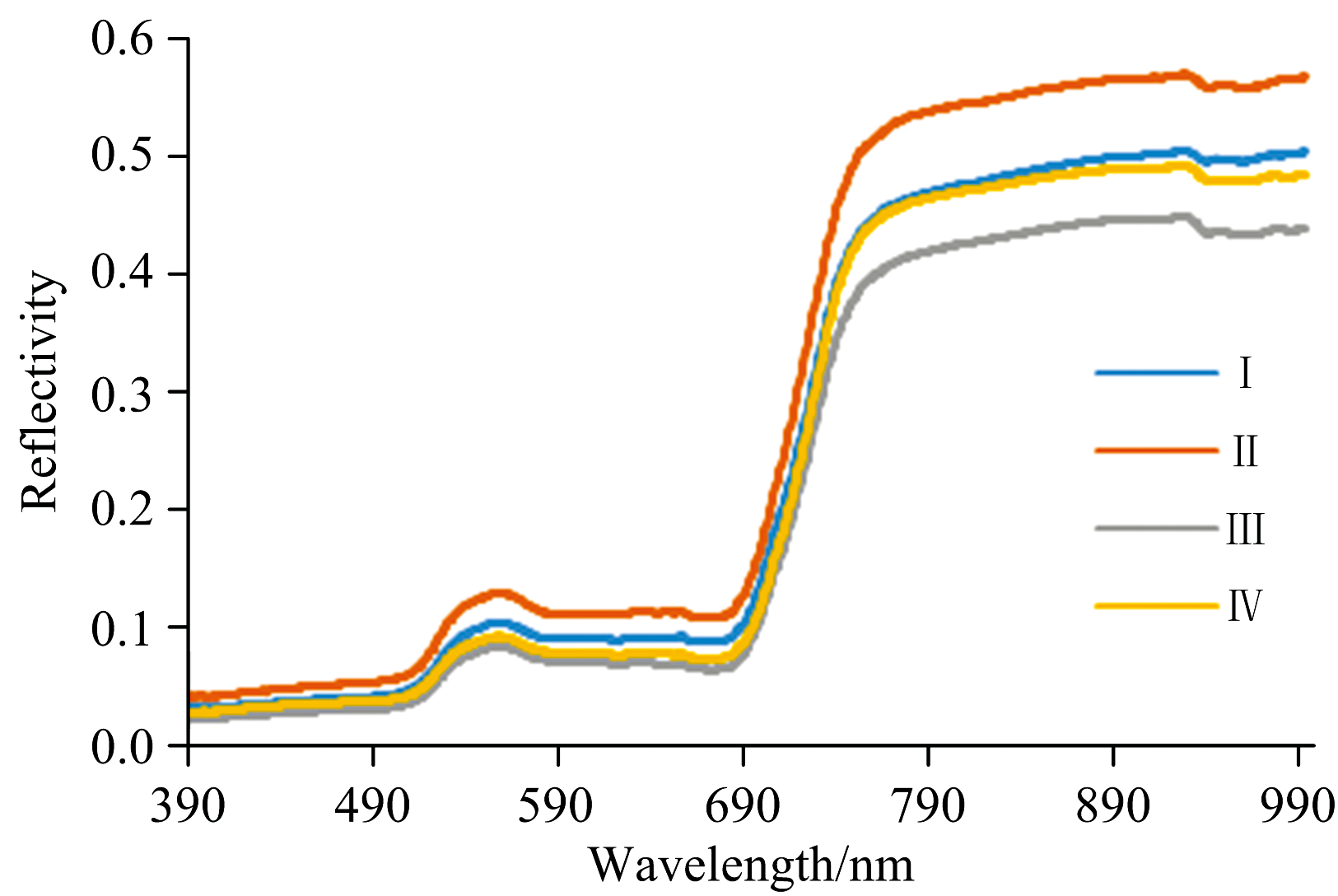 | 图2 向杉木林扩张不同阶段毛竹原光谱反射率Fig.2 Original reflectivity spectra of Phyllostachys edulis at different stages of expansion into Cunninghamia lanceolata forest |
有研究表明, 对原始光谱(R)进行对数、 开方、 倒数、 一阶微分以及二阶微分等数学变换有助于进一步实现光谱信息异化, 从而更好的表征研究对象特征[20]。 毛竹向杉木林扩张不同阶段光谱曲线的差异可以通过光谱数学变换得以增强, 且变换光谱有利于对“ 绿峰” 和“ 红谷” 之间波段的特征研究。 扩张的四个阶段中, 变换光谱反射率变化趋势一致, 以扩张第Ⅱ 阶段为例(图3), 对光谱取倒数(1/R)增加了可见光区反射率量值差异, 对数倒数(1/lgR)变换增加了近红外波段反射率量值差异; 此外, 与前人研究一致[21], 微分变换可以增加光谱的波动性为研究提供更多特征波段, 倒数一阶微分[(1/R)']变换的变化幅度大, 对于“ 绿峰” 和“ 红谷” 处的特征分析更具优势。 但变换形式的差异在不同建模中的贡献也有所不同, 说明不同研究对象对光谱变换形式的敏感形式不同, 不宜简单套用。
 | 图3 向杉木林扩张毛竹光谱变换反射率(以第Ⅱ 阶段为例)Fig.3 Spectral transformations of reflectivity data of Phyllostachys edulis at stage Ⅱ of expansion into Cunninghamia lanceolata forest |
总体来看, 毛竹冠层反射率在不同扩张阶段表现为Ⅱ > Ⅰ > Ⅳ > Ⅲ , 即毛竹扩张边界向杉木林推进会引起冠层光谱的明显变化, 这可能与毛竹种群的生态位转换有关。 毛竹数量较少的扩张前期(第Ⅲ — Ⅳ 阶段)光谱反射率低于毛竹扩张引发“ 林退竹进” 的扩张后期(第Ⅰ — Ⅱ 阶段), 与毛竹扩张前期多通过地下茎潜向生长[22]的机制相符, 随着“ 出笋成竹” 后的快速生长, 毛竹扩张后期冠层反射率升高。 扩张前后即第I阶段和第Ⅳ 阶段反射率差异较小的现象, 可能由于毛竹扩张前尚未显现冠层优势, 而扩张后通过降低种内竞争维持生存优势, 两者表现出相当的冠层高光谱反射率; 而扩张第Ⅱ 阶段与扩张第Ⅲ 阶段反射率的较大差异, 则体现了毛竹种群通过竞争占据优势的生态位转换, 毛竹在第Ⅱ 阶段占据优势生态位, 反射率最高, 而在第Ⅲ 阶段与杉木存在激烈的竞争博弈, 反射率最低。
由毛竹向杉木林扩张不同阶段原始光谱和10种数学变换光谱与毛竹LAI的相关系数最大绝对值确定特征波段, 由表2可见, 毛竹向杉木林扩张不同阶段LAI反演敏感波段在不同阶段表现出一定的集聚现象。 扩张第Ⅰ 阶段的特征波段主要集中在可见光波段的“ 红谷” 附近, 微分变换后的特征波段向“ 绿峰” 偏移; 第Ⅱ 阶段的特征波段主要集中在可见光波段的“ 绿峰” 附近, 微分变换后的特征波段向“ 红谷” 偏移, 说明微分处理可以丰富光谱在“ 绿峰” 和“ 红谷” 之间的变化, 对于更好的区分毛竹在杉木林中的扩张过程具有一定意义; 第Ⅲ 、 Ⅳ 阶段的特征波段主要集中在紫光波段, 表明紫光波段对覆被植物变化的响应或有特征意义[23], 微分变换增加了可见光区特征波段的分布范围, 这两个扩张阶段出现了蓝光波段的特征优势, 尤以第Ⅳ 阶段的蓝光特征更为明显, 也与微分光谱变换会增加特征波段表达相符。 毛竹扩张不同阶段在紫光波段及蓝光波段表现出的光谱特征, 在以往研究中较少被关注, 需进一步揭示其与毛竹和土壤环境理化特征变化之间的联系。
| 表2 毛竹向杉木林扩张不同阶段LAI高光谱反演特征波段 Table 2 Characteristic wavebands of hyperspectral data for LAI inversion at different stages of Phyllostachys edulis expansion to Cunninghamia lanceolata forest |
根据建模集中原始光谱和变换光谱特征波段, 分别与毛竹向杉木林扩张不同阶段的LAI构建五类单因素回归模型, 其中, 二次多项式回归模型是各扩张阶段LAI反演的优势模型, 指数模型次之, 而线性、 幂指数和对数模型表现不佳。 总体来看, 微分处理的光谱变换建模情况表现更优, R'、 R″、 (lgR)'和(1/R)'特征波段所建模型R2更接近于1, RMSE和nRMSE更小; 非微分处理的其他变换光谱仅基于倒数1/R特征波段所建模型表现较好, 但在建模集和验证集间表现差异较大。 从毛竹向杉木林扩张不同阶段来看, 第Ⅰ 阶段二阶微分处理使建模集的决定系数(R2=0.849)最大, 但验证集决定系数较低, 综合来看, 一阶微分处理使建模集和验证集R2分别为0.593和0.174, 更适合第Ⅰ 阶段LAI反演; 第Ⅱ 阶段一阶微分处理使建模集的决定系数(R2=0.863)最大, 但验证集决定系数较低拟合效果不佳, 综合来看, 倒数一阶微分处理使建模集和验证集R2分别为0.597和0.748, 更适合第Ⅱ 阶段LAI反演; 第Ⅲ 阶段二阶微分处理使建模集的决定系数(R2=0.843)最大, 但验证集决定系数较低拟合效果不佳, 综合来看, 对数倒数一阶微分处理使建模集和验证集R2分别为0.735和0.742, 更适合第Ⅲ 阶段LAI反演; 第Ⅳ 阶段倒数一阶微分处理使建模集的决定系数(R2=0.784)最大, 但验证集决定系数较低拟合效果不佳, 综合来看, 一阶微分处理使建模集和验证集R2分别为0.317和0.778, 更适合第Ⅳ 阶段LAI反演(见表3)。
| 表3 基于不同指标的毛竹向杉木林扩张不同阶段LAI反演最优模型 Table 3 Optimal models for LAI inversion at different expansion stages of Phyllostachys edulis to Cunninghamia lanceolata forest based on different indicators |
毛竹向杉木林扩张不同阶段与LAI显著相关的7个植被指数中, YI在扩张第Ⅰ 、 Ⅱ 、 Ⅲ 阶段与LAI显著负相关(相关系数-0.445), 与第Ⅳ 阶段正相关, 从而拉低了整体水平的相关性, 这可能是因为扩张第Ⅳ 阶段毛竹数量较少影响了黄光波段表达毛竹冠层的代表性, 而随着毛竹数量的增加, YI与LAI的相关性比其他植被指数更高, 可见, 黄光指数在毛竹扩张过程中具有代表性; 其他植被指数相关系数由大到小分别为NDVI(0.387)、 mNDSI(0.364)、 GNDVI(0.354)、 NRI(0.351)、 RVI(0.320)、 NVI(0.301), 显著性水平除RVI和NVI在0.05水平上显著相关外, 其余均表现为在0.01水平上显著相关。 根据毛竹向杉木林扩张不同阶段植被指数与LAI的相关系数确定优势植被指数, 再利用建模集植被指数分别与不同扩张阶段LAI构建五类单因素回归模型, 得到不同阶段LAI反演最优模型见表3。 单因素植被指数LAI反演模型以二次多项式模型表现最佳, 但除扩张第Ⅲ 阶段利用NDVI作为优势植被指数构建的模型建模集和验证集R2均大于0.45以外, 其他扩张阶段模型在建模集和验证集拟合效果不佳, 整体来看, 单因素植被指数对毛竹扩张杉木林LAI反演效果不如变换光谱。
YI在毛竹向杉木林扩张第Ⅰ 、 Ⅱ 、 Ⅲ 阶段表现出比其他常用于LAI反演的植被指数更高的相关性, 重申了YI在遥感反演中重要性[10, 24], 表明毛竹扩张LAI的变化会引起黄光波段的特殊变化, 使得YI成为毛竹扩张研究的重要植被指数。 但在不同扩张阶段的模型反演中, YI并未表现出与LAI的高敏感性, 说明黄光波段指数的表征潜力尚未被有效挖掘, 需要进一步通过波段间组合构建新型植被指数, 以发挥黄光波段的更大优势。 NDVI在毛竹向杉木林扩张不同阶段表现出与LAI的高敏感度, 且在模型构建中表现出优于其他植被指数的反演效果, 表明NDVI是毛竹扩张研究的最优植被指数, 但整体来看, 植被指数作为毛竹扩张LAI反演因素的效果并不佳, 显示出LAI反演常用植被指数存在局限性, 需要进一步与气象、 水文等因素结合优化植被指数。
机器学习是当下LAI遥感反演的常用手段, 以往研究在随机森林和神经网络算法方面取得较好模型拟合效果, 但在毛竹向杉木林扩张过程LAI的高光谱反演中效果不理想。 选取的神经网络回归、 贝叶斯线性回归、 决策森林回归和线性回归四类拟合效果相对较好的机器学习中, 只有神经网络在扩张第Ⅰ 阶段R2为0.585, RMSE和nRMSE分别为4.397和22.7%, 拟合度较之其他机器学习方法和其他扩张阶段相对较好。 但与单因素回归模型相比, 机器学习算法在毛竹向杉木林扩张不同阶段LAI反演中有待进一步深耕。
前人研究结果表明, 利用机器学习对植被指数进行多因素回归分析可以在LAI反演方面表现出更大优势[8], 但毛竹向杉木林扩张不同阶段LAI反演最优模型倾向于使用传统回归算法[25], 以基于微分变换光谱的二次多项式和幂指数模型最佳。 微分处理的变换光谱较之经典植被指数表现出更为出色的建模效果, 进一步表明LAI遥感反演需在充分考虑光谱信息的基础上优化植被指数。 神经网络算法建模在毛竹向杉木林扩张第Ⅰ 阶段展示出拟合优势, 表明其更适用于群落结构简单的林分; 扩张第Ⅲ 阶段基于对数倒数一阶微分建模效果最佳, 建模集和验证集R2均在0.7以上, RMSE和nRMSE较小, 表明杉竹各半的林分对研究毛竹向杉木林扩张具有重要价值[4], 在该林分中进行LAI反演效果最好, 利用遥感手段开展毛竹林科学管控宜选竹林各半的林分。
毛竹向杉木林扩张不同阶段在地面高光谱信息中表现出明显差异, 对光谱数据进行微分变换会放大光谱特征表达, 增加特征波段信息, 在不同扩张阶段LAI反演模型构建中表现较佳。 不同扩张阶段在黄光波段、 紫光波段及蓝光波段表现出与毛竹冠层LAI的敏感性对于毛竹扩张的深入研究具有重要意义, 尤其是黄光波段与毛竹扩张的显著相关性是未来需要关注的方向。 扩张第Ⅰ 阶段LAI反演最优模型为利用神经网络算法基于7个植被指数(YI、 NDVI、 mNDSI、 GNDVI、 NRI、 RVI、 NVI)构建的多元回归模型, 验证集R2为0.585, RMSE为4.397, nRMSE为22.7%; 扩张第Ⅱ 阶段LAI反演最优模型为基于倒数一阶微分交换光谱构建的二次多项式回归模型y=-11.96x2-11.43x+2.499 8, 验证集R2为0.748, RMSE为0.370, nRMSE为8.0%; 扩张第Ⅲ 阶段LAI反演最优模型为基于对数倒数一阶微分交换光谱构建的幂指数回归模型y=5.291 4e183.76x, 验证集R2为0.742, RMSE为0.468, nRMSE为9.9%; 扩张第Ⅳ 阶段LAI反演最优模型为基于一阶微分交换光谱构建的幂指数回归模型y=5.092 2e-770.9x, 验证集R2为0.778, RMSE为0.323, nRMSE为6.5%。 综合考虑建模集和验证集LAI反演建模效果, 毛竹向杉木林扩张第Ⅲ 阶段表现最佳, 宜在竹林各半的混交林中开展毛竹扩张科学管控。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|