





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
太湖藻华高发区DOM的组成和时空分布特征
[韩子怡1, 2  , 周永强
, 周永强2 , 耿春女1, * ]
, 周永强]
|
|


作者简介: 韩子怡, 女, 1998年生, 上海应用技术大学生态技术与工程学院硕士研究生 e-mail: hzy15611790201@163.com
随着太湖周边城市社会经济的快速发展, 大量工农业和生活废水进入太湖, 氮磷营养盐富集, 尤其是太湖北部湖区, 藻华频发。 在2015年春、 夏、 秋3个季节对太湖北部藻华高发区24个点进行了6次野外采样, 结合吸收光谱与三维荧光-平行因子分析法, 揭示藻类生消影响下太湖DOM光谱参数的时空分异特征及DOM光谱指标与水质参数的协同变化机制, 为太湖生态系统的治理修复及保障太湖水供给安全提供科学支撑。 在取样时间段(从5月12日—10月20日), 3个湖区水体均经历了一个非常明显的藻华生消过程, 藻华的暴发与外源营养盐的输入密切相关。 夏季期间, 藻类生物量的快速增长显著消耗了水体中的N
With the rapid socioeconomic development in the Lake Taihu basin, a large amount of industrial, agricultural, and domestic wastewater has been discharged into Lake Taihu, and nutrients have been enriched, especially in the northern lake regions of Lake Taihu, where algal blooms occur frequently. In the spring, summer, and fall of 2015, we conducted six field sampling campaigns at 24 sites in northern Lake Taihu′s high-incidence area of algal blooms. Absorbance coupled with three-dimensional fluorescence and parallel factor analysis (3DEEMs-PARAFAC), together with water quality parameters, were used to investigate the spatial and temporal variability of DOM and water quality in Lake Taihu under the influence of algal bloom to provide scientific support for the management and restoration of the Lake Taihu ecosystem and ensure the safety of its water supply. During the sampling period (from May 12 to October 20), the three lake regions experienced significant algal bloom and degradation. The algal blooms were closely related to the input of exogenous nutrients. During the summer, the rapid growth of algal biomass significantly consumed N
湖泊是地表水资源的重要载体和特大、 大中型城市最重要的集中式供水的水源地。 溶解性有机物(dissolved organic matter, DOM)是天然有机质最重要的赋存形态和活跃成分, 主要包含腐殖酸、 脂肪族及芳香烃类等结构复杂的有机物[1]。 外源河流、 地下水、 降水、 工农业及生活污水等的输入、 湖泊内部浮游植物堆积降解、 微生物降解和受风浪扰动的底泥间隙水释放等均会对湖泊DOM来源和组成产生影响。 DOM丰度及来源组成又会直接影响水处理工艺与流程, 大量DOM的赋存会使得水体酸臭刺鼻, 增加水处理成本, 并产生大量致癌消毒副产物。 此外, DOM来源组成还会直接影响微生物与光降解活性[2], DOM经微生物降解和光降解后能释放大量小分子量有机酸和无机碳氮磷营养盐, 为水体中微生物提供大量生源要素的同时又影响湖泊富营养化进程[3]。 因此, 揭示湖泊DOM来源组成特征可为湖泊生态环境修复和维护湖泊水供给安全提供重要保障。
太湖全湖当前处于富营养状态, 北部湖区(尤其是西北竺山湾及西部沿岸带等水域)是太湖营养水平最高的区域, 藻华频发。 近十年来, 夏季在太湖西北沿岸和湖湾静水区, 乃至太湖东北部贡湖湾等静水水域, 蓝藻过量增殖、 堆积和降解, 造成溶氧极低、 大量水生生物死亡及水体发臭水色发黑的“ 湖泛” 现象[4]。 这对太湖水环境治理修复和湖泊供水安全造成严重困扰。 尽管目前已有大量研究围绕太湖蓝藻水华和富营养化开展分析[5, 6, 7], 然而太湖北部藻华动态如何影响DOM来源组成特征并不清晰。 本文通过聚焦太湖北部藻华高发区的DOM时空分异特性, 丰富和发展了现有对于藻类生消过程如何影响DOM组成的相关研究, 并揭示了外源输入与藻类生消如何共同作用于太湖北部DOM库, 进而导致北部部分湖湾及西部入湖沿岸带等区域夏季晴朗静水区出现零星“ 湖泛” 现象, 使得太湖水环境久治难愈, 威胁到湖泊的供水安全。 此外, 研究还利用季节性野外观测数据, 深入分析了藻华高发区DOM的变化特征, 以期为太湖生态系统的治理修复及保障太湖水供给安全提供科学支撑。
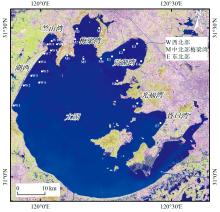
太湖(30° 50'—32° 80'N, 119° 80'—121° 55'E)位于长江三角洲地区的核心位置, 毗邻江苏和浙江两省。 在太湖藻华高发的北部水域(竺山湾、 西部沿岸带、 梅梁湾、 贡湖湾、 光福湾及胥口湾)布设24个采样点(图1, 西部: W1—W11; 北部梅梁湾M1—M6; 东部: E1—E7), 于2015年5月12日和19日、 7月21日和28日、 9月29日和10月20日分别开展6次野外观测。
 | 图1 太湖北部藻华高发区采样点分布示意图Fig.1 Location of sampling sites in the hyper-eutrophic northern half of Lake Taihu |
使用采水器采集表层(0.5 m)处水样, 置于放有冰块的保温箱内避光保存, 采样结束后将样品迅速运回实验室。 分别用0.7 μ m高温450 ℃煅烧过的Whatman GF/F玻璃纤维滤膜和0.22 μ m的Millipore滤膜过滤, 原水和滤液均冷藏于4 ℃的冰箱, 测样前拿出待恢复至室温, 所有样品均在3 d内完成测定。
总氮(TN)和总磷(TP)分别参考过硫酸钾消解紫外分光光度法(GB 11894-89)和钼锑抗分光光度法(GB 11893-89)使用紫外分光光度计(Shimadzu UV-2550, 日本)测定[8]; 叶绿素(Chl-a)的测定需先用90%的热乙醇从0.7 μ m的滤后水中提取叶绿素, 再使用紫外分光光度计(Shimadzu UV-2550, 日本)测定665及750 nm 处的吸光度, 经计算得到Chl-a浓度。
采用分光光度计(Shimadzu UV-2550, 日本)和5cm石英比色皿测定吸收光谱, 水样为0.22 μ m的滤后水, 空白对照为Milli-Q纯水, 以1 nm为间隔测量200~800 nm内DOM的吸光度。 本研究使用波长在350 nm处的吸收系数a350表征DOM的相对丰度。 光谱斜率S275-295由波长范围为275~295 nm内的吸收系数经指数函数拟合得到, S275-295表征DOM芳香性, 其值越小, 表明芳香性越强, DOM的陆源腐殖化程度越高[9]。 紫外吸收系数SUVA254为单位DOC浓度下对应的紫外吸光度[10], 其与DOM的芳香性呈正比, 该值越大, 表明DOM腐殖化及芳香性程度越高, 陆源输入信号越强烈。
三维荧光光谱(three-dimensional excitation-emission matrices, 3DEEMs)能揭示DOM来源组成的信息, 因而近年来被广泛应用于不同类型水体的DOM研究。 平行因子分析法(parallel factor analysis, PARAFAC)可将相互交叠的荧光图谱分离为多个相互独立且具有单个发射波长最大值的荧光峰, 能有效避免传统的“ 寻峰法” 造成解译结果的误判[11]。 采用荧光分光光度计(Hitachi F-7000, 日本)测定三维荧光光谱, 利用MATLAB R2015b的drEEM工具箱进行EEMs矩阵运算和平行因子分析, 选取144个EEMs矩阵, 最终确定4个组分模型能很好地通过对半检验、 随机初始化分析和残差分析, 采用每个荧光组分的最大得分值(Fmax)表征各荧光组分的强度[12]。 腐殖化指数(humification index, HIX)是发射波长为435~480与300~345 nm波段内的荧光强度积分的比值, 两段发射波长对应的激发波长均为254 nm(本研究中由于激发波长间隔为5 nm, 因而该处激发波长为255 nm), 该指数越大, DOM腐殖化程度越高。
使用ArcGIS 10.2绘制采样点位图和插值图, 使用Origin 2021绘制箱线图和散点图, 使用R中的pheatmap包绘制热图并进行层次聚类, 采用IBM SPSS 26对数据进行平均值、 标准差和t检验等相关统计分析, 在本研究中, p< 0.05为显著相关, p< 0.001为极显著相关。
根据点位的空间分布将24个采样点分成3个湖区: 西北部(竺山湾与西部沿岸带)、 中北部梅梁湾、 东北部(贡湖湾、 光福湾与胥口湾), 采用5个指标(Chl-a、 TN、 TP、
 | 图2 太湖北部3个湖区Chl-a、 TN、 TP、 |
进一步探究TN中的无机氮(
3个湖区DOM紫外光谱参数(a350、 SUVA254和S275-295)均存在明显的湖区差异和季节变化(图3)。 表征DOM丰度的a350和芳香性的SUVA254均呈现出由西向东递减的趋势[图3(a, c)]。 S275-295的变化趋势与之相反(图3b), 呈现出由东向西递减的趋势。 S275-295值越小, 芳香性越强, 即陆源类腐殖酸浓度较高。 陆源DOM和氮磷营养物从西北部进入太湖, 顺水流向东南方出湖口扩散, 因此西北湖区DOM库与陆源生活污水及土壤淋溶的输入息息相关, 表现出更高的DOM丰度, 更强的芳香性以及强烈的陆源输入信号。 从季节上看, a350和SUVA254值整体表现为夏季明显高于春秋季[图3(a, c)], S275-295的低值多在夏季出现[图3(b)], 这说明夏季DOM丰度、 芳香性和陆源腐殖化程度均达到最高。
 | 图3 太湖北部3个湖区DOM相关参数的变化特征Fig.3 Variability of DOM-related parameters in the northern half of Lake Taihu |
虽然西北湖区和中北湖区的藻华情况都较为严重, 但其紫外参数存在着差异性。 夏季西北部和中北部的Chl-a浓度均值分别为118和188 μ g· L-1, 中北部梅梁湾的藻华在夏季更严重[图2(a)]。 与5月19日相比, 7月21日西北湖区和中北湖区的a350分别增加0.23和0.78倍, SUVA254分别增加0.01和0.16倍, S275-295分别降低1%和11%, 这表明伴随着藻华发生, 梅梁湾湖区DOM丰度、 芳香性和陆源输入信号增加更快, 一方面是因为藻源性DOM提高了梅梁湾湖区DOM丰度[16], 另一方面是因为夏季降水增加, 水体连通性增强, 径流携带更多的陆源DOM进入湖体, 影响范围从西北部入湖口延伸到中北部梅梁湾湖区。
对6次野外观察的144个3D-EEM进行PARAFAC分析, 解析出4种荧光组分(图4), 分别是2种类蛋白质组分(C1、 C3)和2种类腐殖质组分(C2、 C4)。 组分C1(Ex/Em ≤ 230(285)/340 nm)为类色氨酸组分, 表征生物降解或生活污水来源; 组分C3(Ex/Em=280/316 nm)为类酪氨酸, 表征藻降解输入; 组分C2(Ex/Em=240/388 nm)表征微生物作用产生的类腐殖质[17]; 组分C4(Ex/Em≤ 230(250、 360)/468 nm)则是典型的陆源性类腐殖质[18]。
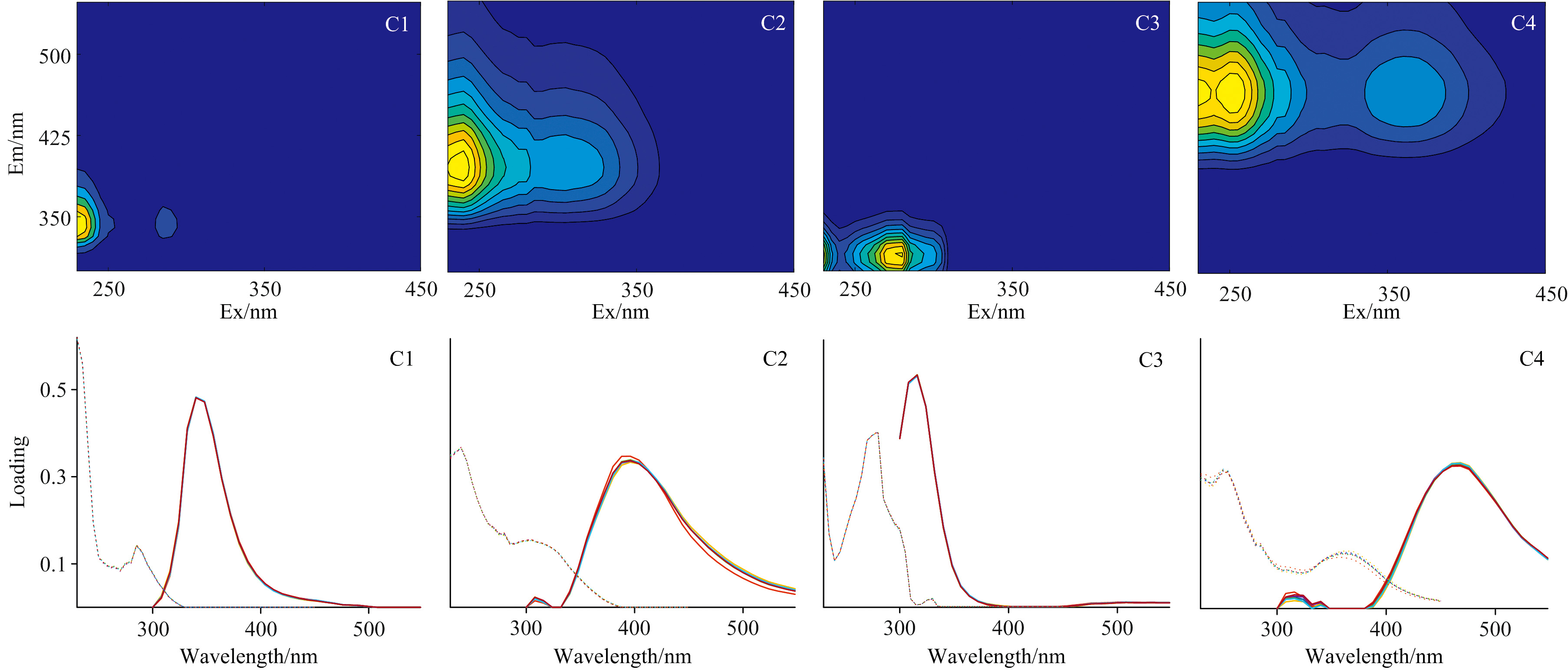 | 图4 荧光组分(C1—C4)的三维荧光光谱和对半检验图Fig.4 Fluorescent shapes of the four components derived from PARAFAC-modeling and the half-split validation results |
藻华生消的过程中, 4个荧光组分展现出明显的时空特异性。 类蛋白质组分(C1和C3)在时间上表现出相似的变化模式: 从春季到夏季, C1和C3明显降低, 到秋季又回归到春季水平[图5(a—f, m—r)]。 这一现象的原因是, 夏季藻华期间, 依附蓝藻的藻附生菌和浮游细菌大量增殖, 微生物活动会优先利用类蛋白质中的小分子物质, 导致夏季类色氨酸C1和类酪氨酸C3降低, 进入秋季, 藻华消退, 藻类衰亡降解产生大量类蛋白质[19], 外加微生物活动的减少, 使类蛋白质的荧光强度得以累积至初始水平。
 | 图5 荧光组分C1—C4的6次野外调查期间的空间分布Fig.5 Spatial distribution of fluorescence intensities of the four PARAFAC components C1—C4 |
研究进一步发现C1和C3的空间分布存在着季节差异, 在春季, C1荧光强度自西向东递减, C3则是自东向西递减; 到了夏季, C1的空间分布不再遵循明确规律, 仅在部分区域出现高值, 而C3的空间分布仍保持春季的趋势; 到了秋季, C1和C3均呈现出自西向东的趋势。 这表明C1的空间分布受到人类活动和藻华的双重影响, 特别是西北部生活污水的输入使C1在春秋季呈现出西高东低的趋势, 而在夏季, 由于藻类在各个区域的繁殖和活动强度不一, C1的空间分布无明显的规律; C3主要受到湖区内的生物特性和藻华的影响, 西北部和中北部处多为蓝藻繁殖, 而贡湖湾多为黄藻, 胥口湾处则多为水草[20], 春夏季东北部湖区的黄藻及水生植物活动较强, 秋季西北部和中北部湖区大量蓝藻衰亡, 不同区域藻类的繁殖和衰亡造成了C3独特的空间分布。
两种类腐殖质(C2和C4)的荧光强度展现出一致的时空变化[图5(g—l, s—x)], 荧光强度在空间上均自西向东递减, 在时间上均表现为夏秋季较高。 这一现象主要是由于西北部和中北部湖区高人口密度和强烈的人类活动, 导致大量陆源类腐殖质C4通过径流输入湖泊[21]。 夏季雨水丰沛带来更多的陆地凋落物和土壤有机质, 进一步加剧了湖泊的腐殖化程度。 同时, 藻华的暴发促进了微生物的活动, 微生物活动利用类蛋白质生成了类腐殖质, 导致微生物源的类腐殖质C2增加。 进入秋季, 降水减少使得湖内水体更新速度下降, 死亡的藻类大量堆聚在水面, 形成了避光环境, 阻碍了类腐殖质的光降解进程[22], 从而在水体中累积了更多的类腐殖质。 藻华生消的过程中, 类蛋白质组分始终是DOM的主要组成部分, 藻华生消影响了不同DOM组分的相对贡献量, 使更多的内源性DOM参与到湖泊碳循环过程中。
太湖北部湖区Chl-a与DOM吸收系数a350呈极显著正相关[p< 0.001, 图6(a)], 表明藻类生物量的增多会提高DOM丰度。 藻类生长代谢和死亡降解都会释放大量内源性DOM, 因此藻类浮游植物生长降解是太湖DOM的重要来源之一[16]。 Chl-a与S275-295呈极显著负相关[p< 0.001, 图6(b)], 且与SUVA254和HIX均呈极显著正相关[p< 0.001, 图6(c, d)], 表明外源输送的大量高芳香性陆源DOM为浮游藻类生长提供了重要碳源, 是藻华发生的物质基础。 此外, Chl-a与C1、 C3均呈显著负相关[p< 0.05, 图6(e, g)], 与C2、 C4均呈极显著正相关[p< 0.001, 图6(f, h)], 进一步表明藻华生消过程中会影响DOM的组成。
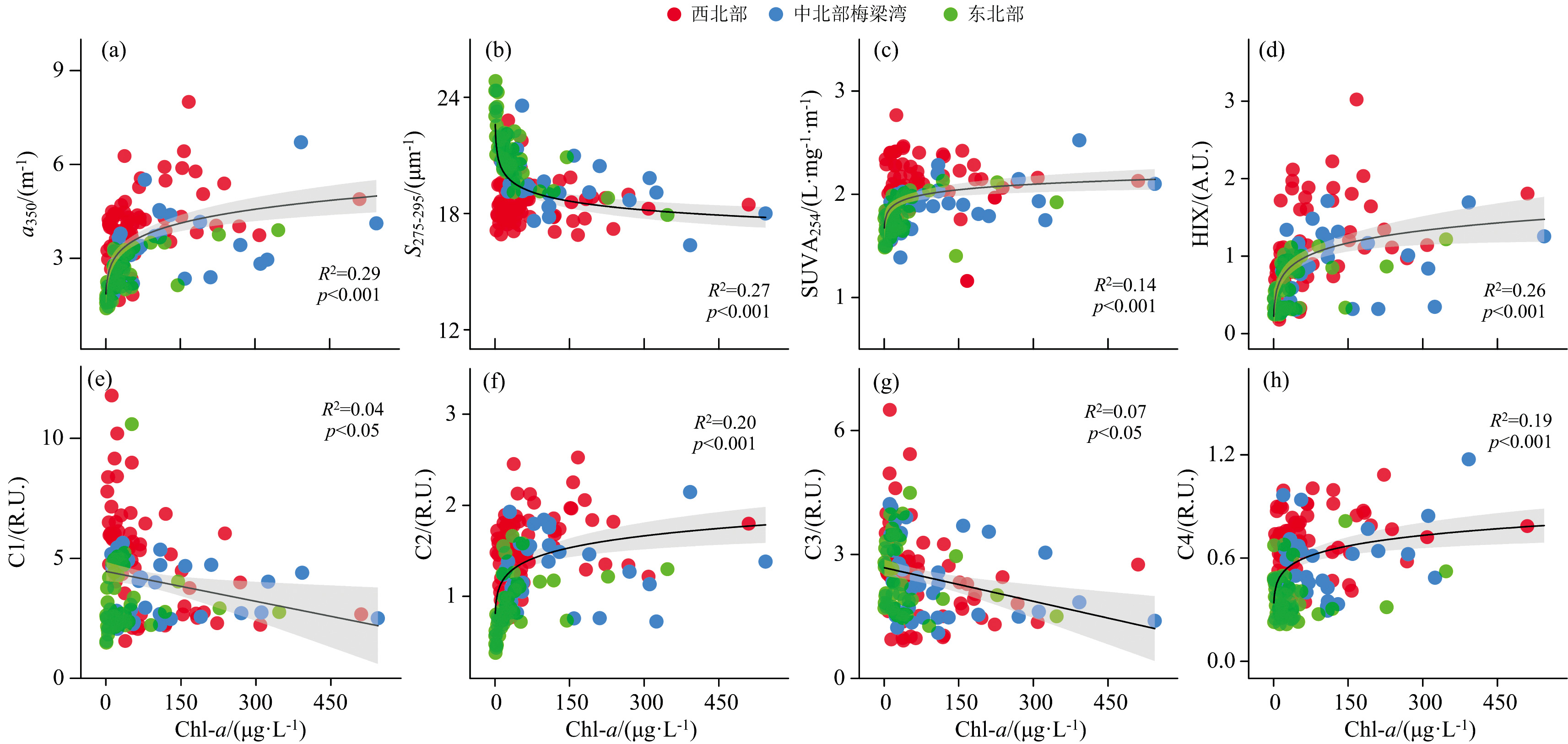 | 图6 太湖北部3个湖区a350、 S275-295、 SUVA254、 HIX、 C1、 C2、 C3、 C4和Chl-a之间相关性分析Fig.6 Relationships between Chl-a and a350, S275-295, SUVA254, HIX, C1, C2, C3, C4 in the hyper-eutrophic northern half of Lake Taihu |
根据DOM光谱参数的组成(C1、 C2、 C3、 C4、 HIX、 a350、 SUVA254、 S275-295), 对24个采样点的6次野外观察数据分别进行层次聚类(图7)。 从时间角度分析, 春季, 西北部湖区的8个点位因高DOM丰度(a350)和高外源输入(C1、 C4和HIX高, S275-295低)而聚成一类。 夏季, 西北部湖区所有点位、 中北部部分点位以及东北部的一些点位因高DOM丰度(a350)和高外源输入(C4和HIX高, S275-295低)而聚成一类, 这与图3的研究结果一致, 表明夏季更多陆源DOM进入湖体, 藻华发生影响范围由西部湖区延伸到东北部湖区。 进入秋季, 东北部湖区所有点位和中北部一些采样点因低陆源输入信号(S275-295高)聚成一类, 同时西北部和中北部的一些点位因强烈的微生物活动(C2高)聚成一类, 这表明秋季水体更新速度变慢, 湖区之间的联系较夏季降低, 西北部和中北部部分区域受到了藻华降解的影响, 微生物活动剧烈。
 | 图7 基于6次野外观察的DOM参数对取样点进行分组的树状热图Fig.7 Heatmap with dendrograms of sampling sites clustered by the optical composition of DOM during the six field sampling campaigns |
从区域特征分析, 西北部湖区S275-295始终保持着较低值, 此湖区始终受到外源输入的影响; 中北部湖区的8个参数在时间上显示出最大的变异性, 藻类生消极大程度上影响了此湖区的DOM组成及结构; 东北部部分参数(a350、 S275-295)表现出较少的变化, 此湖区受藻华动态和陆源输入的影响较小。 因此, 面对太湖北部的藻华问题, 必须实施区域差异化的治理策略: 西北部需削减外源营养流入, 中北部梅梁湾应加强监测与生态恢复, 而东北部则应聚焦生态多样性的维护以提升自净能力。 同时, 考虑到气候变暖导致的湖温升高和蓝藻提前爆发的问题, 可以利用湖泊的反硝化作用控制蓝藻水华, 例如种植沉水植物, 利用沉水植物与微生物的联合作用提高反硝化速率[23], 在水华暴发前降低湖泊内的氮浓度, 在控磷的基础上协同控氮, 以遏制太湖蓝藻水华的发生。
本研究仅捕捉了六次时间节点下的藻华高发区水体DOM组成及结构的动态变化, 在整体时间维度上仍存在着局限性, 可能无法完全捕捉到太湖水体DOM组成及藻华状况的最新变化。 同时, 本研究主要依赖于传统的水质监测和样品分析方法, 这些方法在时空上的分辨率可能限制了对快速变化事件的捕捉能力。 因此, 未来研究应采用长期连续监测和高分辨率遥感技术, 结合多学科方法, 深入分析人类活动和自然因素的交互作用, 并开发新的生态修复策略, 提升太湖水质管理的科学性和预见性。
(1)太湖北部3个湖区均经历了一个非常明显的藻华生消过程, 藻华的暴发与外源营养盐的输入密切相关。 夏季期间, 藻类生物量的快速增长显著消耗了水体中的
(2)太湖北部水体的营养负荷、 藻类生物量以及DOM丰度、 芳香性和陆源腐殖化程度均呈现出由西向东递减的趋势, 外源输入影响了太湖北部水体的营养水平和DOM组成及结构, 陆源DOM也为浮游藻类生长提供了重要碳源, 使微生物活性强的内源DOM(C2和C3)更多地参与湖泊碳循环过程。
(3)三个湖区中, 藻华暴发显著提升了中北部梅梁湾的DOM丰度, 东部湖区受外源输入和藻华生消影响最小, 其生物特征与其他地区存在显著差异性。 因此, 这三个湖区的藻华治理需因地制宜, 西北部应减少外源输入, 梅梁湾处应加强藻华监测和生态修复, 东北部需要保护和恢复生态多样性, 增强自然净化能力。
致谢: 感谢张成英、 薛静琛、 夏忠等在野外采样及室内实验过程中提供的帮助。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|