{kind=link}
{kind=link}
{kind=link}
{kind=link}
根区增温对玉米幼苗主要代谢物傅里叶红外光谱特性及叶绿素含量的影响
[夏镇卿 , 司雷勇, 金岩, 扶亚芳, 王奇, 路海东
, 司雷勇, 金岩, 扶亚芳, 王奇, 路海东* ]
, 司雷勇, 金岩, 扶亚芳, 王奇, 路海东]
|
|


作者简介: 夏镇卿, 1996年生, 西北农林科技大学农学院硕士研究生 e-mail: 2716555460@qq.com
为深入研究根区温度对玉米幼苗主要代谢物以及叶绿素含量的影响, 试验设置常温(24 ℃)、 中温(30 ℃)和高温(36 ℃)三个根区温度水平, 以SD609(保绿型)和SD902(衰老型)两个不同抗逆型玉米品种为样品, 采用傅里叶变换红外光谱仪(Tensor Ⅱ, 德国布鲁克)测定玉米幼苗根、 茎、 叶三个器官的光谱特征。 结果表明: 不同根区温度处理对玉米幼苗各个器官有机物含量有显著影响, 但影响程度不同, 大体表现为根>茎>叶; 玉米幼苗根、 茎、 叶在3 330, 2 927, 1 639, 1 515, 1 350, 1 250和1 055 cm-1等波数处透射率均受根区温度的影响, 且在根区中温条件下各器官上述波数处透射率较低, 根区高温条件下透射率较高, 在1 055 cm-1处各处理差异最为明显; 不同抗逆型品种在根区增温条件下FTIR光谱特征有较大差异, 抗逆性较差的衰老型品种(SD902)在不同根区温度处理下FTIR光谱特征差异较大, 叶绿素含量受根区增温影响较大, 而抗逆性较好的保绿型品种(SD609)不同处理间差异较小, 叶绿素含量受根区增温影响较小; 根区温度对不同玉米品种根系活力及叶绿素含量影响均表现为中温(30 ℃)>对照(24 ℃)>高温(36 ℃)。 因此, 适当提高根区温度可以显著提高玉米幼苗根系活力、 叶绿素、 碳水化合物尤其是多糖含量, 以及茎秆和叶片中蛋白质、 核酸含量, 但温度过高会使多糖、 脂类、 叶绿素含量以及根系活力迅速下降; 与SD609相比, SD902对根区温度反应较为敏感, 在根区高温条件下, 玉米内源物质合成受阻, 而SD609能保持相对较高的多糖及叶绿素含量, 对根区高温条件有相对较好的适应性。 根区温度对玉米幼苗根系作用最大, 可通过影响根系吸收、 物质合成及物质转运功能, 进而影响茎秆、 叶片有机物以及叶绿素含量。
In order to study the effects of root zone temperature on the main metabolites and chlorophyll content of maize seedlings, the spectral characteristics and chlorophyll content of two maize cultivars under three root zone temperature levels were determined by Fourier transform infrared spectroscopy and SPAD-502 chlorophyll meter. The results showed that different root zone temperature treatment had significant effects on the organic matter content of various organs of maize seedlings, but the degree of influence was different, and the general realization was root>stem>leaf. The transmission of wave number of maize seedlings in 3 330, 2 927, 1 639, 1 515, 1 350, 1 250 and 1 055 cm-1 was affected by the temperature of the root zone. The peak value was lower under medium temperature conditions and higher under the high temperature condition of the root zone, and the difference was most obvious at 1 055 cm-1. The FTIR infrared spectral characteristics of different varieties in the root zone temperature range are quite different, and SD902 has a large difference, while SD609 has a small difference. The temperature of root zone had significant effects on the root activity and chlorophyll content of different maize varieties, and both showed medium temperature (30 ℃)>control (24 ℃)>high temperature (36 ℃). In summary, appropriate increase of root zone temperature can significantly increase the content of chlorophyll, root activity, carbohydrates, especially polysaccharides in corn seedlings, as well as protein and nucleic acid content in stems and leaves, but excessive temperature will rapidly decrease the content of polysaccharides, root activity, lipids and chlorophyll. The increase in root zone temperature has a significant effect on the seedling metabolites of SD902 and SD609 corn varieties, and the SD902 is more sensitive to the temperature response in the root zone, and the SD609 has better heat resistance. The root zone temperature had the greatest effect on the roots of maize seedlings, and by effecting the roots absorption, material synthesis and material transport functions, and then affect the stalk, leaf organic matter and chlorophyll content.
近年来全球温度持续升高, 温度变化对作物生长发育的影响, 已成为一个不容忽视的问题[1]。 随着全球气候变暖趋势的不断加剧, 作物根区土壤温度也在发生着显著变化[2]。 玉米作为我国最为重要的粮食饲料作物之一, 保证其产量品质, 对保障我国粮食安全具有重要意义。 温度变化对玉米生长发育的影响已成为当下研究的热点, 但目前的研究多集中于冠层温度对玉米的影响[3], 而根区温度对玉米生长发育尤其是对玉米幼苗期植株中有机大分子及叶绿素含量的影响方面鲜有报道。
根区温度对玉米生长发育的影响主要有根系及冠层形态、 根系衰老、 养分水分吸收速率以及一些生理性状等, 根区温度过高过低都会影响玉米生长[4]。 前人研究表明, 根区温度超过35 ℃时玉米根系生长速率会降低, 当低于4.5 ℃时, 玉米根系停止生长[5], 同时过高的根区温度会使玉米根系及茎秆粗度变细, 对玉米光合特性、 根系活力、 内源激素及酶活性都有显著影响[4]。 探寻温度变化下不同抗逆型玉米品种的植株代谢特征对指导未来玉米育种和安全生产具有重要意义。
傅里叶变换红外光谱(FTIR)是一种基于有机物分子中极性键振动来分析物质组成的技术, 可以对样品进行定性定量分析, 通过吸收峰的高低可以准确反映出植株中蛋白质、 碳水化合物、 脂类以及核酸含量的高低, 具有灵敏度高、 操作安全性高、 测试方法简单、 测试时间短、 样品用量少等优点[6]。 周丽平等[7]应用FTIR研究了腐殖酸对玉米植株中蛋白质、 碳水化合物、 核酸等有机大分子含量的影响。 欧全宏等利用FTIR研究了水稻、 玉米、 蚕豆正常叶和稻瘟病叶、 玉米和蚕豆锈病叶的红外光谱特性, 发现两者之间的碳水化合物和蛋白质的含量存在差异。 但鲜有研究应用FTIR方法来检测根区增温条件下玉米幼苗根、 茎、 叶中有机大分子含量变化。 采用水培的方法, 通过改变水温来模拟土壤温度变化, 解决了传统方法土壤水分对试验结果的干扰, 以及传统方法增温不均匀不稳定的问题。 应用FTIR及SPAD仪测定不同温度处理下玉米幼苗根、 茎、 叶中有机大分子含量变化和叶绿素含量, 研究根区温度对玉米生长的作用机理, 为全球气候变暖土壤温度升高背景下玉米绿色安全生产提供理论依据。
供试玉米为抗逆性较好的保绿型品种陕单609(SD609)和抗逆性较差的衰老型品种陕单902(SD902), 由农业部西北旱区玉米生物学与遗传育种重点实验室提供。 选择饱满一致的玉米种子经70%的酒精消毒5 min, 放入蒸馏水中浸种12 h, 再放入发芽盒中催芽2 d后选取生长一致的幼苗, 定植于装有10L的1/2霍格兰营养液的塑料盒(40 cm× 30 cm× 12 cm)中, 每盒24株, 每个品种3个处理, 每个处理3盒, 放于24 ℃恒温培养室继续培养, 待幼苗长至一叶一心时, 在塑料盒中放入变频恒温加热棒(中山市日宝公司生产, 功率100 W, 控温精度± 0.2 ℃)进行精确根区增温处理, 处理如下: 根区中温处理(MT, 恒温加热设备使水温保持在30 ℃)、 根区高温处理(HT, 恒温加热设备使水温保持在36 ℃)、 对照(CK, 根区不增温, 温度保持在24 ℃, 此温度依据陕西关中地区大田春玉米苗期土壤温度设定)。
1.2.1 根系活力测定
处理8 d后, 采用TTC法进行根系活力测定[8]。
1.2.2 叶绿素含量
处理8 d后, 利用SPAD-502叶绿素测量仪测定玉米幼苗最后一片展开叶的叶绿素含量。
1.2.3 傅里叶变换红外光谱分析
处理8 d后, 将玉米幼苗根、 茎、 叶烘干, 磨碎然后过0.2 mm网筛, 将样品与溴化钾(KBr)按1∶ 100的比例混合后, 放入玛瑙研钵中研磨混匀, 进行压片, 采用傅里叶变换红外光谱仪(Tensor Ⅱ , 德国布鲁克)测定玉米幼苗根、 茎、 叶的光谱特征(扫描范围4 000400 cm-1, 分辨率2 cm-1, 扫描次数64次)。
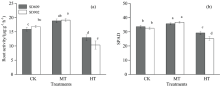
随着根区温度升高, 两个品种根系活力呈先上升后下降的趋势[图1(a)], MT处理下SD609与SD902根系活力分别较对照升高18.72%和13.61%(p< 0.05), 在CK和MT处理下SD902根系活力要略高于SD609; HT处理下两品种根系活力迅速下降, 与CK相比SD609根系活力下降18.50%(p< 0.05), 而SD902根系活力下降38.53%(p< 0.01), 在36 ℃根区高温处理下SD902根系活力要比SD609低20.03%(p< 0.05)。 可见, 根区高温条件下SD609更能维持较高的根系活力。
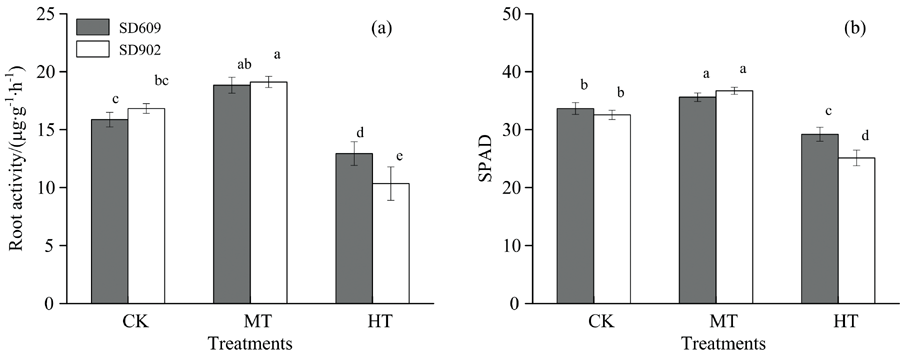 | 图1 不同根区温度处理下玉米幼苗根系活力和SPAD值Fig.1 Root activity and SPAD value of maize seedling under different root zone temperature |
在不同根区温度处理下, SPAD值差异显著[图1(b)], 且都是中温处理SPAD值最高, 高温处理SPAD值最低, 两者之间差异达到极显著(p< 0.01)。 与对照相比, SD609中温处理SPAD值升高了5.79%(p< 0.05), 高温处理降低了13.22%(p< 0.05)。 SD902中温处理与对照相比SPAD值升高了12.75%(p< 0.05), 高温处理降低了22.89%(p< 0.05)。 可见, 保绿型玉米品种SD609在温度变化下叶绿素合成比较稳定, 高温胁迫下能够维持相对较高的叶绿素含量; 而衰老型玉米品种SD902在温度变化下叶绿素合成变化幅度较大, 玉米幼苗期适当的增加根区温度有利于叶绿素含量的显著升高, 但根区温度过高会显著降低玉米幼苗期叶片叶绿素含量。
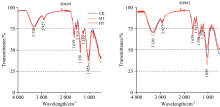
不同根区温度处理下玉米幼苗主要官能团及其FTIR的特征波长见表1。 在不同根区温度处理下, 玉米幼苗根系的傅里叶变换红外光谱的特征峰位置无显著差异, 但不同品种及不同处理各个官能团对应特征波长的透射率有显著差异(图2)。 两个品种根区高温处理在3 330, 2 927和1 055 cm-1处透射率高于中温处理和对照。 由于3 330 cm-1附近的吸收峰是由碳水化合物中的饱和O— H以及蛋白质、 氨基酸、 核酸的N— H伸缩振动的叠加效应产生(表1), 所以根区高温处理在此处的透射率高于其他处理, 说明36℃根区高温对玉米幼苗期根系生长有一定抑制作用, 而中温处理此处透射率较其他处理显著降低, 其中以SD902根区中温处理下降幅度最明显, 而SD609随着根区温度的变化差异相对较小。 说明衰老型玉米SD902根系对根区温度反应较为敏感, 适当地提高根区温度有利于玉米幼苗的生长发育。 2 927 cm-1附近的吸收峰是脂类化合物中饱和— CH2中的C— H伸缩振动产生(表1), 此峰主要来自细胞内各种膜及细胞壁的成分, SD609根区中温处理此处透射率略低于根区高温处理和对照, 处理间差异不大, SD902根区中温处理与对照无显著差异, 而根区高温处理此处透射率要显著高于对照, 说明36 ℃根区高温胁迫对SD902幼苗期根系细胞壁、 细胞膜及其他的一些膜结构有破坏作用。 1 055 cm-1附近吸收峰由多糖中的C— O和C— C伸缩振动产生(表1), 根区增温对玉米幼苗根系中多糖的影响较为明显, 从图3可以看出, 随着根区温度的升高, 各个根系多糖含量呈现先增加后减少的趋势, 且36 ℃处理下SD902多糖含量最低。 说明根区增温有利于玉米幼苗期根系多糖积累, 但温度过高反而会使多糖含量迅速降低, 而衰老型玉米品种SD902根系多糖含量受根区温度影响较大。
| 表1 玉米植株主要有机物、 官能团及其对应的FTIR特征波长 Table 1 Characteristic wavelengths of main functional groups in maize |
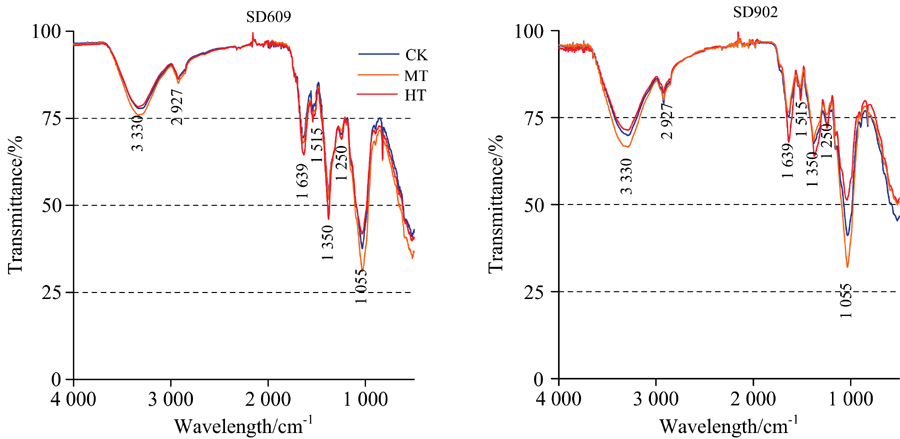 | 图2 不同根区温度处理下玉米幼苗根系的傅里叶变换红外光谱(FTIR)谱图Fig.2 FTIR spectra of maize seedling root |
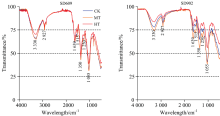
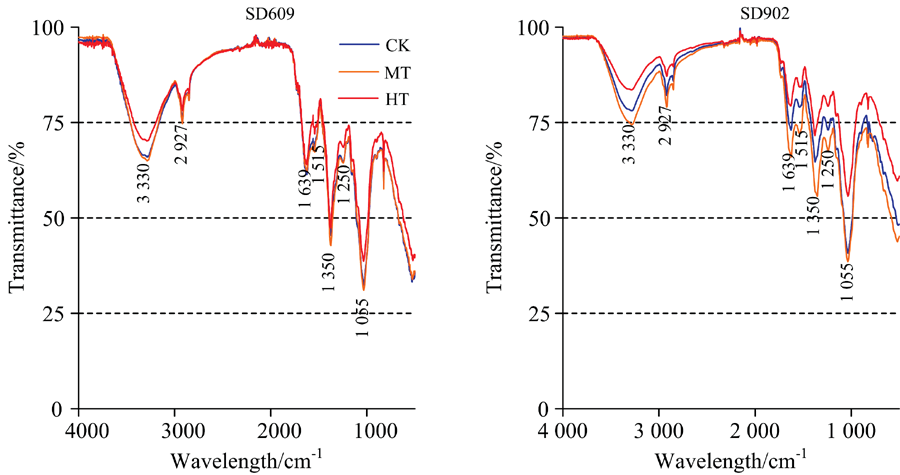 | 图3 不同根区温度处理下玉米幼苗茎秆的傅里叶变换红外光谱(FTIR)谱图Fig.3 FTIR spectra of maize seedling stalk |
另外, 两品种根区高温处理后在1 639, 1 515, 1 350和1 250 cm-1处透射率有不同程度的降低。 1 750~1 500 cm-1波数中的1 639 cm-1吸收峰是酰胺化合物中的氨基酸、 多肽类物质中N— H变形振动产生, 另外1 350 cm-1附近的吸收峰为蛋白质中— CH2和— CH3的弯曲振动(表1), 相比于中温处理和对照, SD902和SD609根区高温处理在1 750~1 500和1 296~1 352 cm-1处透射率有较为明显的降低, 表明了根区高温胁迫下玉米幼苗根系蛋白质、 氨基酸、 多肽含量是升高的, 这可能与高温胁迫下氨基酸的积累效应与植物的抗逆性有关, 同时由于植物的抗逆机制, 在1 250 cm-1附近核酸降低, 以此来合成大量的酶和蛋白质来应对根区高温胁迫, 相对于衰老型品种SD902, 保绿型品种SD609的根系抗高温机制明显增强。
在根区高温胁迫下, 各品种玉米幼苗茎的红外光谱在3 330, 2 927, 1 639, 1 515, 1 350, 1 250和1 055 cm-1处透射率有不同程度的升高, 根区增温处理对不同品种茎秆发育影响程度不同, SD902各处理间差异较大, SD609各处理在3 330和1 055 cm-1处有显著差异, 其他特征波长处差异较小, 与根系对根区高温反应不同, 各品种茎秆在根区高温胁迫下蛋白质、 氨基酸、 多肽及核酸含量也有所降低, SD902降低幅度最为明显。 同时, 各品种根区中温胁迫茎秆中碳水化合物、 蛋白质、 核酸以及多糖含量要高于对照。 由此可知, 根区温度的变化对玉米幼苗期茎秆中主要有机物含量有显著影响, 茎秆中的蛋白质、 氨基酸、 多肽、 多糖、 核酸含量随着温度的升高呈现先升高后降低的趋势, 在36 ℃高温处理下各种物质含量最低, 适当提高根区温度有利于玉米幼苗期茎秆的发育, 但高温胁迫显著阻碍了其玉米茎秆中的内源物质合成, 造成茎秆发育不良。 衰老型品种SD902茎秆发育受根区高温影响明显大于保绿型品种SD609, 相对于衰老型品种, 在根区高温胁迫下, 保绿型品种茎秆中具有相对稳定的蛋白质、 核酸以及多糖合成。
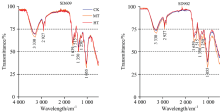
不同根区温度处理的玉米幼苗期叶片红外光谱谱图存在一定差异(图4), 但与根系、 茎秆相比, 不同处理间差异较小; 两品种在3 330, 2 927, 1 639, 1 515, 1 350, 1 250和1 055 cm-1波数特征峰处透射率总体上表现为HT> CK> MT, CK与HT之间差异不显著, SD609 MT处理在3 330和1 055 cm-1处透射率较低, SD902 MT处理在3 330, 1 639, 1 515, 1 350, 1 250和1 055 cm-1处透射率有显著降低。 说明本试验根区温度对于玉米幼苗期叶片主要有机物含量的影响要比对根系和茎秆的影响小, 根区温度对SD609叶片多糖含量以及SD902叶片多糖、 蛋白质、 核酸含量影响较大, 过高根区温度不利于植物叶片中多糖、 蛋白质、 核酸的合成。
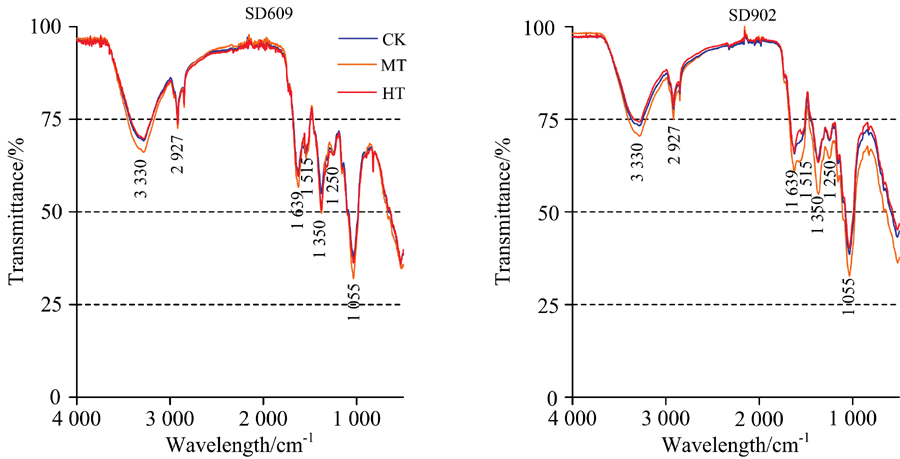 | 图4 不同根区温度处理下玉米幼苗叶片的傅里叶变换红外光谱(FTIR)谱图Fig.4 FTIR spectra of maize seedling leaf |
氮和镁是叶绿素的基本组成成分, 铁、 锰、 锌、 铜等元素在叶绿素合成中有重要作用[11], 这些叶绿素合成需要的营养元素主要通过植物根系吸收, 根系生长发育及吸收能力可能是造成不同根温条件下玉米幼苗期叶绿素差异的主要原因, 根区温度可以通过调控玉米幼苗根系活力来间接影响叶绿素含量。
实验发现, 根区温度影响了玉米幼苗期根、 茎、 叶的傅里叶红外光谱特征, 适当提高根区温度有利于玉米幼苗期根、 茎、 叶碳水化合物尤其是多糖含量的提高, 以及叶片茎秆中蛋白质、 核糖含量的升高。 而当根区温度过高时, 玉米幼苗期根、 茎、 叶碳水化合物、 脂类、 含量下降, 以碳水化合物含量下降幅度最为明显, 这与前人研究发现的玉米在根区高温胁迫下干物质积累量快速下降这一现象一致[5]; 同时, 茎秆和叶片中蛋白质、 氨基酸、 多肽、 核酸含量也有所下降。 根区温度对玉米幼苗期各个器官的影响程度表现为根系> 茎秆> 叶片。 衰老型品种SD902对根区温度反应较为敏感, 根区中温处理下SD902长势要略优于SD609; 但在36 ℃根区高温胁迫下SD902根、 茎、 叶中多糖含量明显下降, 同时根系中脂类含量也出现较为明显的下降, 表明保绿型品种SD609比衰老型品种SD902具有更好的耐热性。
根区高温处理下, 蛋白质、 氨基酸、 多肽以及核酸含量有所升高, 这主要是由于植物的抗逆机制。 有研究表明, 高温胁迫可以促进抗逆性相关的酶以及蛋白质尤其是可溶性蛋白的生成[12], 本试验中玉米根系可能通过核酸转录翻译出大量氨基酸、 蛋白质来形成相关的酶和可溶性蛋白以抵御根区高温胁迫。 根区高温条件下玉米根系脂类化合物含量降低明显, 这可能是过高的温度破坏了根系的生物膜系统。 而玉米茎秆和叶片中蛋白质、 脂类以及核酸含量的变化可能主要与不同处理间根系的吸收及输送功能有较大的关系。 根区增温处理下玉米幼苗期各个器官碳水化合物含量变化最明显且呈现先升高后降低的趋势, 可能有以下几点原因: 首先植物生长所需糖类主要由叶片光合作用产生并运送到各个器官, 根区增温处理通过影响根系的吸收功能, 从而影响了玉米叶绿素的含量, 从源头上导致了不同处理玉米各个器官碳水化合物的差异; 其次不同器官不同处理之间碳水化合物含量差异的显著性不同, 根系差异最大, 而叶片差异最小, 这可能是因为根区土壤温度影响了玉米输导组织的运输效率。 有研究表明, 高温胁迫可以影响植物维管束的发育[13], 本试验中根区高温胁迫可能使玉米幼苗各部分维管束发育不良, 从而导致叶片生成的光合产物运输受阻, 另外, 根区增温可能会影响植物尤其是根系的呼吸作用, 温度升高呼吸作用也会升高, 碳水化合物消耗速率也会加快[14], 这可能也是造成不同处理间碳水化合物差异的一个重要因素。
玉米幼苗根系活力和叶绿素含量与土壤温度显著相关(p< 0.05), 适当提高根区温度有利于植物根系活力及叶绿素含量的升高, 但根区温度过高会降低玉米幼苗期的根系活力及叶绿素含量, 研究发现, 衰老型玉米品种SD902在根区高温胁迫下叶绿素含量下降最明显, 耐热性较差; 而保绿型品种SD609的耐热性相对较好, 根区高温胁迫下叶绿素含量下降幅度较小。 两品种根区增温处理下根系活力变化与叶绿素含量变化趋势相似, 在高温胁迫下, 相对于衰老型品种, 保绿型品种具有更好的根系吸收能力。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|