


{kind=link}
{kind=link}
{kind=link}
大豆叶片呼吸与植被指数和叶片性状的关系
[王瑾1, 2  , 陈书涛
, 陈书涛1, 2, * , 丁司丞1, 2 , 姚雪雯1, 2 , 张苗苗1, 2 , 胡正华2 ]
, 陈书涛, 丁司丞|
|


作者简介: 王 瑾, 女, 1997年生, 南京信息工程大学应用气象学院硕士研究生 e-mail: 17863205133@163.com
为研究大豆叶片呼吸与植被指数和叶片性状的关系, 设置田间试验, 观测不同生长阶段的大豆顶1叶、 顶2叶、 顶3叶叶片呼吸及呼吸系数, 并观测归一化植被指数(NDVI)、 差值植被指数(DVI)、 比值植被指数(RVI)、 增强植被指数(EVI)、 光化学植被指数(PRI)、 红边叶绿素指数(RECI)6种高光谱植被指数以及叶绿素相对含量(SPAD)值、 叶片鲜重、 叶片干重、 含水量、 叶面积、 比叶面积、 氮含量。 结果表明: 大豆单片叶片呼吸及呼吸系数存在明显的季节变化规律, 顶1叶、 顶2叶、 顶3叶单片叶片呼吸季节平均值分别为(0.157±0.019), (0.162±0.014)和(0.142±0.010) mg·d-1, 其呼吸系数季节平均值分别为(0.638±0.072), (0.678±0.082)和(0.642±0.076) mg·g-1·d-1, 顶1叶、 顶2叶、 顶3叶季节平均单片叶片呼吸及呼吸系数均无显著( p>0.05)差异。 不同植被指数的季节动态之间存在差异, RVI, EVI, PRI和RECI在生长季中期的数值相对更大, RVI, EVI, PRI和RECI表现为单峰曲线的变化规律。 除大豆生长初期外, 随着叶片位置下降, SPAD值、 鲜重、 干重、 叶面积均逐渐下降。 叶片含水量随着叶片生长呈现出下降的规律。 单片叶片呼吸与气温、 PRI存在极显著( p<0.01)相关关系, 与RECI、 氮含量存在显著( p<0.05)相关关系, 基于这4个因子的模型可模拟单片叶片呼吸60.4%的季节变异。 呼吸系数与干重、 比叶面积存在极显著( p<0.01)相关关系, 与气温、 SPAD值存在显著( p<0.05)相关关系, 基于这4个因子的模型可模拟叶片呼吸系数72.4%的季节变异。 本研究显示, 大豆叶片呼吸与高光谱植被指数和叶片性状存在内在联系, 采用高光谱植被指数可有效模拟不同位置叶片呼吸及呼吸系数的季节变异。
A field experiment was performed to investigate the relationships between the leaf respiration of soybean and vegetation indexes and leaf characteristics. The respiration and respiration coefficient of the first, second and third leaves from the top at the different growth stages were measured. The normalized difference vegetation index (NDVI), difference vegetation index (DVI), ratio index (RVI), enhanced vegetation index (EVI), photochemical reflectance index (PRI) and red edge chlorophyll indexes (RECI) were obtained from the hyperspectral method as well as the leaf characteristics of chlorophyll SPAD value, fresh mass, dried mass, water content, leaf area, specific leaf area and nitrogen content were also determined. The results showed that the respiration of a single leaf and respiration coefficient had obvious seasonal patterns. The seasonal mean respiration of the single first, second or third leaf from the top was (0.157±0.019), (0.162±0.014) and (0.142±0.010) mg·d-1, respectively. The seasonal mean respiration coefficient of the first, second or third leaf from the top was (0.638±0.072), (0.678±0.082), (0.642±0.076) mg·g-1·d-1, respectively. There were no significant ( p>0.05) differences in the seasonal mean leaf respiration and respiration coefficient between the first, second or third leaf from the top. There were significant ( p<0.05) differences in the seasonal patterns between the different vegetation indexes. The relatively high RVI, EVI, PRI and RECI appeared mid-growth stages. The seasonal patterns of RVI, EVI, PRI and RECI showed a single unimodal curve. The SPAD value, fresh mass, dried mass and leaf area decreased with the decrease in leaf position except for at the beginning growth stages. The leaf water content decreased with the growth of leaf growth. The respiration of a single leaf was highly significantly ( p<0.01) correlated with the RECI and nitrogen content. The respiration of a single leaf was significantly ( p<0.05) correlated with the air temperature and PRI. A model based on these four factors explained 60.4% of the variation in the respiration of a single leaf. The respiration coefficient was highly significantly ( p<0.01) correlated with thedried mass and specific leaf area. The respiration coefficient was significantly ( p<0.01) correlated with the air temperature and SPAD. A model based on these four factors explained 72.4% of the variation in the respiration coefficient. The present study showed that the leaf respiration of soybean could be linked with the hyperspectral vegetation indexes and the leaf characteristics. The seasonal variations in the leaf respiration and leaf respiration coefficient in the different positions could be effectively modeled with the hyperspectral vegetation indexes.
温室气体排放导致的全球变暖是人们关注的重要环境问题。 CO2是最主要的温室气体, 大气中CO2浓度升高与全球碳循环有关[1]。 绿色植物叶片在通过光合作用固定CO2的同时还通过呼吸作用向大气中排放C
不同植物的叶片呼吸存在差异, 且同一种植物在不同生长阶段的叶片呼吸也不同, 叶片呼吸与一系列叶片生长指标和叶片性状有关[4]。 以往研究表明, 具有不同代谢活性的不同类型植物的叶片呼吸存在差异, 这种差异与叶片基因型、 大小、 形状、 质量等因素有关[5, 6]。 同一种植物在不同生长环境和生长阶段下的叶片呼吸与比叶面积有关, 不同植物类型的叶片呼吸系数均与比叶面积呈现出线性回归关系[7]。 研究表明, 叶片呼吸还与叶片氮含量表现为线性回归关系[8], 氮含量高低对叶片呼吸的影响可能体现了蛋白质周转过程和活性对生命维持活动的影响程度大小, 两者间的这种关系不仅体现在不同植物类型上, 而且体现在同一种植物不同位置的叶片上[9, 10]。 然而关于这方面的研究还相对较少, 与自然生态系统相比, 不同作物不同生长阶段叶片的呼吸也存在差异。 利用高光谱技术测得的植被指数可反映作物的某些生理生长特征(如: 绿度、 叶面积指数)[11, 12], 作为叶片生长状况指标的叶片呼吸与高光谱植被指数可能存在联系。 然而关于作物叶片呼吸与植被指数关系的研究非常鲜见, 而基于植被指数和叶片性状的作物叶片呼吸的模拟模型则更加缺乏。
本研究观测大豆不同生长阶段的叶片呼吸及呼吸系数, 并观测高光谱植被指数和叶片叶绿素含量、 叶重、 叶面积、 比叶面积、 氮含量等叶片性状, 分析叶片呼吸及呼吸系数与植被指数和叶片性状的关系, 以期为更准确地估算作物呼吸提供基础数据和理论支撑。
在南京信息工程大学农业气象试验站(32.21° N, 118.70° E)进行田间试验。 试验地多年平均降水量为1 100 mm, 多年平均气温为15.6 ℃, 供试土壤为黄棕壤。 0~20 cm耕层土壤质地为壤质黏土, 土壤田间持水量25.6%, 容重1.54 g· cm-3, pH(H2O)为6.30, 有机碳含量8.91, 全氮含量0.81 g· kg-1。
于2020年06月至2020年10月进行田间试验, 在3个面积分别为2.5 m× 2.5 m的小区种植大豆(Glycine max L.), 大豆品种为八月白。 于2020.06.09施用基肥(复合肥51.3 g· m-2), 复合肥N:P2O5:K2O之比分别为15%:15%:15%。 于2020年06月09日播种大豆, 大豆三叶期、 分枝期、 开花期、 结荚期分别为2020年06月16日, 2020年06月23日, 2020年07月24日和2020年08月17日。
于2020年06月30日, 2020年07月10日, 2020年08月03日, 2020年08月20日, 2020年09月14日和2020年10月05日期间采用红外CO2气体分析仪(SoilBox-FMS, Sable systems公司, USA)测定每个小区中各6片顶1叶(植株最顶端叶片)、 顶2叶(从顶端数第2片叶片)、 顶3叶(从顶端数第3片叶片)的叶片呼吸,
在测定大豆叶片呼吸的当天中午(此时光照条件较好), 采用便携式地物光谱仪(Fieldspec 4 Standardard-Res, ASD公司,
| 表1 植被指数的计算公式 Table 1 Functions to compute vegetation indexes |
将不同日期烘干后的大豆顶1叶、 顶2叶、 顶3叶磨碎, 过100目筛, 采用凯氏定氮法测定大豆叶片全氮含量[15]。
采用单因素方差分析判断顶1叶、 顶2叶、 顶3叶季节平均单叶片呼吸及呼吸系数的差异显著性。 以Pearson相关分析判断单叶片呼吸及叶片呼吸系数与NDVI、 DVI、 RVI、 EVI、 PRI、 RECI、 SPAD值、 鲜重、 干重、 含水量、 叶面积、 比叶面积、 氮含量的关系。 建立基于高光谱植被指数与叶片性状的单叶片呼吸和呼吸系数多元回归方程, 以方差膨胀因子(VIF)来判断各潜在影响变量的共线性, 当某因素VIF< 10时, 可作为多元非线性回归的自变量。 以观测的单叶片呼吸及呼吸系数与模拟的单叶片呼吸及呼吸系数进行回归模拟,
单片叶片呼吸存在明显的季节变化规律, 最高的顶1叶单叶片呼吸出现在2020年08月03日, 最低的顶3叶单叶片呼吸出现在2020年10月05日[图1(a)]。 叶片呼吸系数总体呈逐渐降低的季节变化规律, 大豆生长后期的叶片呼吸系数低于生长前期, 顶1叶、 顶2叶最大呼吸系数均出现在2020年06月30日[图1(b)]。 就季节平均值而言, 顶1叶、 顶2叶、 顶3叶单叶片呼吸无显著差异(p> 0.05), 其呼吸系数也无显著差异(p> 0.05)。
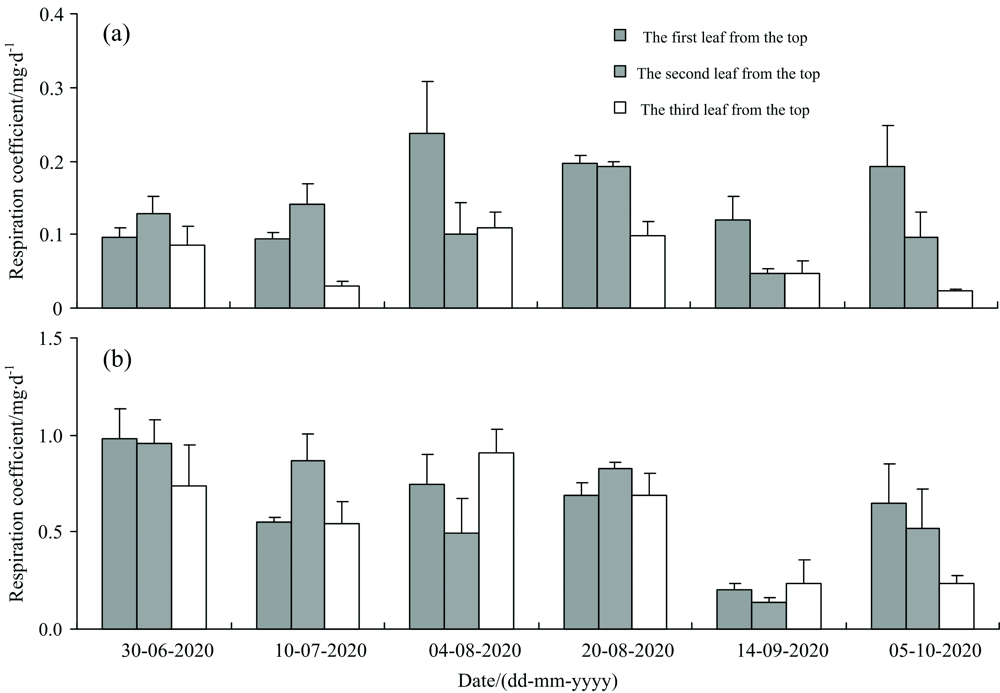 | 图1 单片叶片呼吸及叶片呼吸系数的季节变化 (a): 单叶片呼吸; (b): 叶片呼吸系数Fig.1 Seasonal changes in the respiration of a single leaf and leaf respiration coefficient (a): Reasonal of a single leaf; (b): Leaf respriration coefficient |
由图2(a— f)可见, 不同高光谱植被指数的季节动态之间存在差异, NDVI在不同时期的差异性相对其他植被指数而言更小一些, RVI, EVI, PRI和RECI在生长季中期的数值相对更大, 而生长初期和生长末期的数值相对更小, 表现为单峰曲线的变化规律, 例如EVI和PRI的最大值分别出现在2020年08月20日和2020年08月03日。 与呼吸系数类似, 2020年10月05日的EVI, PRI和RECI值高于2020年09月14日的值, 这说明作物生长后期不同叶片之间的性状仍存在较大差异。
 | 图2 (a) NDVI, (b) DVI, (c) RVI, |
叶片SPAD值变异范围为(34.58± 2.05)~(52.23± 1.62), 除2020年06月30日的生长初期外, 其余观测日顶3叶SPAD值总体上最低。 单片叶片鲜重的变异范围为(0.19± 0.05)~(1.41± 0.17) g,
Pearson相关分析表明, 单叶片呼吸与气温存在极显著(p=0.009)相关关系, 单叶片呼吸与PRI存在极显著(p=0.009)相关关系, 与RECI(p=0.024)和氮含量存在显著(p=0.015)相关关系(表2)。 呼吸系数与气温存在显著(p=0.017)相关关系, 呼吸系数与鲜重、 干重、 比叶面积存在极显著(p< 0.001)相关关系, 呼吸系数与SPAD值存在显著(p=0.022)相关关系。 有的高光谱植被指数之间存在极显著(p< 0.001)相关关系, 例如NDVI与RVI, NDVI与RECI, DVI与EVI等。 有的叶片性状之间存在显著(p< 0.05)或极显著(p< 0.001)相关关系, 例如叶片面积与SPAD值之间存在显著(p=0.032)相关关系, 与鲜重、 干重之间存在极显著(p< 0.001)相关关系。
| 表2 叶片呼吸与气温、 NDVI、 DVI、 RVI、 EVI、 PRI、 RECI、 SPAD值、 鲜重、 干重、 含水量、 叶面积、 比叶面积、 氮含量的Pearson相关分析 Table 2 Pearson's correlation between the leaf respiration and air temperature, NDVI, DVI, RVI, EVI, PRI and RECI, SPAD value, fresh mass, dried mass, water content, area, specific leaf area and nitrogen content of leaf |
多元回归分析表明, 基于气温(AT)、 PRI、 RECI、 氮含量(N)的回归方程可模拟单叶片呼吸(R)60.4%的变异[见式(1)], 基于气温(AT)、 SPAD值、 叶片质量(DM)、 比叶面积(SLA)的回归方程可模拟叶片呼吸系数(Rc)72.4%的变异[见式(2)]。 对观测的单叶片呼吸和呼吸系数与模拟的单叶片呼吸和呼吸系数进行的回归分析表明, 观测值与模拟值之间具有极显著的回归关系, 其斜率分别为0.996 9和1.196 1, 非常接近1:1的比例线[图3(a, b)]。 描述单叶片呼吸和呼吸系数模型模拟效果的RMSE, MAE, AIC和BIC均较低, 而ME均较高。
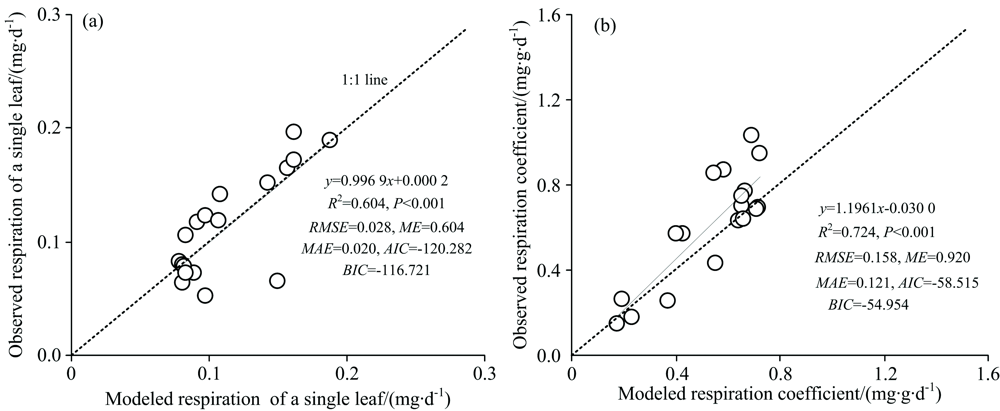 | 图3 观测的单片叶片呼吸及呼吸系数与模拟的单片叶片呼吸与呼吸系数的关系 (a): 单叶片呼吸; (b): 叶片呼吸系数Fig.3 Relationships between the observedrespiration of a single leaf and respiration coefficient and the modeled respiration of a single leaf and respiration coefficient (a): Reasonal of a single leaf; (b): Leaf respriration coefficient |
作物叶片呼吸是重要的叶片生物学指标, 反映了叶片维持其自身生命活动的能力, 叶片呼吸的底物来自于光合作用, 因而与光合作用有关的因素如叶片生长速率、 生物量大小、 氮含量、 叶片性状均影响叶片呼吸[6]。 植被指数通过冠层反射率计算得到, 而反射率反映了植物色素含量、 元素含量、 冠层密度、 生物量等指标, 这些指标也是影响叶片呼吸的潜在变量。 叶片呼吸与植被指数具有内在联系(表2)。
本研究中, 测定大豆叶片呼吸速率的时间与光谱测定时间(中午)较为接近, 忽略大豆叶片呼吸速率日变化的原因一方面是本研究中主要考虑中午光谱数据的准确性, 另一方面是主要考虑大豆叶片呼吸在不同生育期及叶片位置的差异, 且大豆叶片呼吸在不同生育期及叶片位置的差异与高光谱指数对应。 以往研究表明顶1叶至顶3叶是监测作物物质含量的最佳叶位, 其在各个生育期的代表性比下部叶片更好[20, 21], 由于叶片呼吸作用与氮素含量有关[9], 本研究选择顶1叶、 顶2叶、 顶3叶作为测定呼吸作用的叶片。 以往研究表明, 热带雨林冠层顶部叶片呼吸高于下部叶片呼吸, 且上部叶片干重大于下部叶片, 而上部叶片比叶面积小于下部叶片[22, 23]。 本研究中, 就整个生长季平均而言, 大豆顶1叶、 顶2叶、 顶3叶单叶片呼吸和呼吸系数无显著(p> 0.05)差异, 而生长中后期大豆顶1叶单叶片呼吸大于顶2叶和顶3叶。 造成这种差异的原因可能与大豆不同冠层高度光照和热带雨林不同冠层高度光照的差异有关, 本研究中大豆生长初期植株高度对光照的影响相对较小, 这可能使得大豆不同高度植株得到的光照相差不大, 而在大豆生长中后期随着植株高度对光所受照的影响增大, 其单叶片呼吸也产生了差异。
研究显示了单叶片呼吸与高光谱植被指数的显著相关性。 高光谱植被指数反映了作物生长状况, 而叶片呼吸也与作物生长过程有关[24, 25]。 研究中6个植被指数在作物生长初期和生长末期均相对较低, 而在作物生长中期(即旺盛生长阶段)均相对较高, 特别是PRI和RECI的单峰曲线季节变化模式相对更为明显。 植被指数是作物生长发育动态, 如色素、 结构性指标的综合性作用结果, 其差异性体现了作物生长发育动态变化。 在作物生长中期, 叶片生长较快, 为后期的作物结实储存营养物质; 在作物生长末期, 叶片活性减弱, 色素含量发生变化, 而植被指数可反映这些不同生长阶段的差异。 单叶片呼吸与高光谱植被指数的联系表明叶片呼吸可通过高光谱植被指数模拟。 尽管高光谱植被指数随作物生长阶段的不同而存在差异, 但PRI和RECI均与单叶片呼吸存在显著(p< 0.05)相关关系。 以往研究表明, EVI比NDVI可更好地反映作物冠层结构, 包含EVI的模拟生态系统呼吸的模型比包含NDVI的模型可更好地解释生态系统呼吸的变异[26]。 本研究表明, 基于气温、 PRI、 RECI、 氮含量的模型可较好地解释单叶片呼吸。 研究中叶片呼吸系数与高光谱植被指数无显著相关性的原因主要在于高光谱植被指数反映的是作物冠层总体状况, 而呼吸系数表征的是单位质量叶片的呼吸强度, 其主要与作物叶片性状有关(表2)。 SPAD值与光合作用有关, 而光合作用为叶片呼吸供给碳源, SPAD值与呼吸系数的关系间接反映了叶绿素与光合作用的联系。 Reich等[27, 28]的研究表明, 植物叶片呼吸系数与其比叶面积和氮含量之间均存在极显著(p< 0.01)线性回归关系。 孙文娟等[29, 30]的研究表明, 冬小麦和水稻地上部分呼吸系数与氮含量呈极显著(p< 0.01)线性回归关系, 向土壤中添加氮元素可提高植株氮含量进而提高地上部分呼吸系数。 氮是叶片蛋白质的重要组成元素, 对维持蛋白质活性和转运起重要作用[31], 呼吸系数与氮含量的关系体现了作物叶片呼吸与蛋白质周转和代谢等生命活动有关[32]。 表2表明, 具有更大质量和比叶面积的大豆具有相对更高的呼吸活性(即呼吸系数更高), 具有这些特征的叶片需要维持与碳固定有关的更高的代谢活性[33], 其呼吸系数也更高。 比叶面积反映了叶片生长效率和维持生命活动的效率, 也与呼吸系数有关[34]。 大豆单叶片呼吸和呼吸效率与叶片性状之间的相关性体现了叶片不同性状和生理活性的内在关联性。
在模拟单叶片呼吸和呼吸系数时, 需采用不同的高光谱植被指数和叶片性状指标。 叶片呼吸与植被指数具有显著相关性(表2), 特别是单叶片呼吸与PRI的相关系数r达到0.597(p=0.009), 在模拟叶片呼吸时, 在温度和叶片性状指标基础上加入植被指数因子可较好地解释叶片呼吸的季节变化, 分别如图3(a, b)所示。 本研究中基于地面观测的高光谱植被指数和微观尺度的叶片性状指标为基于高空和卫星遥感的作物叶片呼吸的反演提供了可能, 进而可对更大范围的叶片呼吸进行估算。 今后需对更多种作物叶片呼吸与高光谱植被指数和叶片性状的关系进行研究, 以期得到更具广泛性的结果。
(1)大豆单片叶片呼吸和呼吸系数均存在明显的季节变化规律, 但顶1叶、 顶2叶、 顶3叶的单片叶片呼吸和呼吸系数之间均无显著差异。
(2)不同高光谱植被指数的季节变化规律不同。 除大豆生长初期外, 随着叶片位置下降, SPAD值、 鲜重、 干重、 叶面积均逐渐下降。
(3)基于气温、 PRI、 RECI、 氮含量的模型可模拟单片叶片呼吸60.4%的季节性变异, 而基于气温、 SPAD值、 叶片干重、 比叶面积的模型可模拟叶片呼吸系数72.4%的季节性变异。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|