



{kind=link}
{kind=link}
{kind=link}
{kind=link}
金盏菊原生质体对土壤铅/镉响应机制的FTIR, 2D-IR和XPS证据
[范春辉1, 2  , 郑金焕
, 郑金焕3 , 刘宏鑫1 ]
, 郑金焕|
|


作者简介: 范春辉, 1982年生, 沈阳师范大学生命科学学院副教授 e-mail: fanchunhui@synu.edu.cn
土壤修复是“十四五”期间国家重点支持的环保领域, 是实现社会可持续发展的重要保障。 与其他方法相比, 植物修复技术整体优势突出, 对于土壤重金属的去除净化更为有效。 原生质体是植物细胞代谢活动的重要场所, 相对于细胞壁而言, 原生质体对重金属胁迫的生理响应同样强烈。 现阶段, 同类植物修复机制研究多从分子生物学层面切入; 本研究则从谱学角度展开, 初步探究植物原生质体对土壤重金属的反馈信号。 以代表性的菊科植物金盏菊为研究对象, 通过Pb/Cd胁迫盆栽实验获取金盏菊样本, 差速冷冻离心法得到金盏菊原生质体。 引入Tessier连续提取-原子吸收光谱法(AAS)揭示胁迫强度与Pb/Cd赋存形态的内在关联, 结合X射线衍射光谱(XRD)、 傅里叶变换红外光谱(FTIR)、 二维相关红外光谱(2D-IR)和X射线光电子能谱(XPS)识别金盏菊原生质体对Pb/Cd响应的谱学表现。 结果表明: 金盏菊原生质体可交换态Pb/Cd比例不高, 胁迫强度对不同形态Cd含量影响很小。 XRD图谱最强信号出现在31.7°(NaCl晶体), 同时检测出Pb盐 [Pb5(PO4)3Cl] 和Cd盐(CdS)特征峰。 FTIR图谱的3 510 cm-1附近强吸收带源于—OH伸缩振动, 胁迫过程导致峰形杂乱、 峰位偏移; 2D-IR结果说明Pb/Cd优先与金盏菊原生质体—OH和C=O结合。 从XPS图谱可以看出, 反应前后原生质体C, O元素结合能有异。 C(1 s)结合能略有增加, 说明C原子参与了配位反应; O(1 s)峰位有所偏转, 暗示含O基团对Pb/Cd的结合包含多种途径。 新出现的Pb(4 f)峰源于π电子-Pb的交互作用; 胁迫浓度增加导致Cd(3 d)结合能升高, 表明Cd具有明显失电子倾向。 相关结果可以与前期获得的Pb/Cd/金盏菊细胞壁结合特性互为补充, 对于完善同领域的深度和广度、 构建植物修复理论和技术体系意义重大。
Soil remediation is the key issue supported by the national government in the 14th Five-Year Plan for Economic and Social Development of P. R. China, and also important for socially sustainable development. Phytoremediation might remove metals from the soil more effectively compared to other methods. As the dominant location of metabolism in a plant cell, protoplast can get respond greatly to the reaction signal to heavy metals, similar to that of the plant cell wall. Currently, it is also significant to further reveal the remediation mechanism and response signal between plant protoplast and heavy metals during the phytoremediation process, with spectral analysis methods than molecular diagnostic techniques. Calendula officinalis was obtained via pot experiments under lead/cadmium contaminated soil, and the protoplast sample was derived with differential centrifugation technology. Tessier's Sequential Extraction Procedure-Atomic Absorption Spectrometry (AAS) was applied to study lead/cadmium speciation variation. The X-ray diffraction pattern (XRD), Fourier transform infrared spectroscopy (FTIR), two-dimensional infrared spectroscopy (2D-IR) and X-ray photoelectron spectroscopy (XPS) were used to investigate the spectral characteristics of protoplast under different lead/cadmium contents. The results show: the exchangeable contents of lead/cadmium in protoplast are relatively low, and the contents of cadmium with different speciation change little under different lead/cadmium concentrations in soil. The representative peak located at 31.7° in the XRD pattern, the evidence of sodium chloride crystal, and peaks of Pb5(PO4)3Cl and CdS could also be easily found. The dominant absorbance band in FTIR spectra appears at 3 510 cm-1, caused by the vibration of —OH. The peak becomes complicated and moves to some extent, and functional groups of —OH and C=O show a preferential combination to lead/cadmium, indicated from 2D-IR spectra. The elemental binding energies of C and O vary during the reaction in XPS spectra. The binding energy of C(1 s) increases, and C atoms involve in complexation. The moving peak of O(1 s) suggests the reaction nature might be complex. The new peak of Pb(4 f) is caused by the interaction of π electron and lead ions. The binding energy of Cd(3 d) increases at higher lead/cadmium contents, and cadmium would lose electrons during the reaction. The achievements, together with the results related to metals and Calendula officinalis cell wall obtained previously, will play an important role in the theory and technological improvement of phytoremediation.
土壤污染是环境领域的“ 三大战役” 之一, 已发展成全球普遍存在的生态环境问题。 土壤污染能够扰乱生态系统的正常运转, 严重威胁人类生存和农业可持续发展[1]。 作为一个处于快速工业化进程中的发展中国家, 我国的土壤污染总体形势相当严峻。 2016年, 《中华人民共和国土壤污染防治行动计划》(简称“ 土十条” )正式印发实施, 这为我国土壤污染防治工作提供了技术标准和行动纲领。 随着国家“ 十四五” 规划的实施和“ 二○三五” 远景目标建议的提出, 土壤污染修复将迎来新的历史机遇和挑战。
植物修复法(phytoremediation)通过植物自身吸收、 挥发、 根滤、 降解、 稳定化等行为完成土壤污染物的去除净化, 是现阶段及未来土壤修复主流技术之一[2]。 自1983年Chaney提出植物修复技术至今, 国内外学者对其进行了大量探索性研究, 奠定了植物修复技术理论体系的坚实基础。 金盏菊是最常见的城市草本花卉之一, 兼具药用、 食用和装饰等多维价值。 近年研究初步表明: 金盏菊对土壤重金属也有一定去除效果, 可能会成为土壤修复的潜在植物源[3]。 目前, 相关研究尚处于前期探索阶段, 无法系统提供有说服力的基础数据和参考证据, 对于金盏菊吸收土壤重金属的谱学机制研究亟待加强。
前期笔者建立并优化了金盏菊体内Pb/Cd的检测方法, 明确了金盏菊细胞壁与Pb/Cd的体外接触特性[4]。 原生质体是植物细胞的功能性组分, 其对土壤Pb/Cd的体内响应和结合贡献同等重要。 通过Pb/Cd胁迫土壤盆栽体系获取金盏菊植株样本, 引入差速冷冻离心法提取金盏菊原生质体, 借助XRD, FTIR, 2D-IR和XPS剖析Pb/Cd与金盏菊原生质体结合的反馈信号, 力图从光谱学层面为后续相关研究提供理论支持。
偏振塞曼原子吸收光谱仪(Z-2000, Hitachi), X射线衍射仪(D/max2200PC, Rigaku), 傅里叶变换红外光谱仪(VECTOR-22, Bruker), X射线光电子能谱仪(ESCALAB 250 Xi, Thermo Scientific)。 实验用水为桶装纯净水, KBr为光谱纯, 其余化学试剂为优级纯。
向土壤中均匀喷洒250 mL Pb/Cd混合溶液, 充分搅拌使土壤润湿并混合均匀, 即得Pb/Cd复合污染土壤。 土壤染毒过程包含A(CK), B, C和D四个处理, Pb/Cd浓度分别为0/0, 200/10, 400/20和600/30 mg· kg-1。 盆栽周期为2016年4月6日至5月20日; 盆栽容器为圆形塑料盆(盆底直径12 cm, 盆顶直径16 cm, 盆高14 cm), 每盆装有染毒土壤(1.0± 0.01) kg; 依次向其中施加尿素(0.4 g· 盆-1)、 磷酸二氢钠(0.2 g· 盆-1)和氯化钾(0.3 g· 盆-1)三种底肥, 每盆播入20粒金盏菊种子, 维持土壤含水量为田间持水量的70%, 于室外天然光照下培养, 之后收获金盏菊植株用于后续使用。
采用差速冷冻离心法分离金盏菊细胞组分, 所有操作均在4 ℃低温下进行。 称取(1.0± 0.01) g金盏菊鲜样于预冷研钵中, 加入经4 ℃预冷的10 mL提取剂 [0.25 mol· L-1蔗糖, 50 mmol· L-1 Tris-HCl缓冲液(pH 7.5)] 研磨。 将处理后的匀浆分两次倒入6 mL离心管中离心(10 000 r· min-1, 10 min), 下层沉淀即为细胞壁及其破碎残渣组分, 其余产品为金盏菊原生质体。 另取一份金盏菊样品装入消煮管中, 向其中依次加入20 mL HNO3和4 mL HClO4, 常温静置12 h后消解样品, 直至白烟基本散尽。 将冷却后的消解液转移至容量瓶, 定容后摇匀检测Pb/Cd赋存形态。 上述所有实验重复三次。
采用Tessier连续提取-AAS法识别实验样品Pb/Cd赋存形态, 其中AAS为空气-乙炔火焰, 乙炔气体压力160 kPa, Pb/Cd测定波长分别为283.3和228.8 nm。 XRD分析为Cu Kα 辐射源, 扫描角度10° ~70° , 步宽0.02° 。 FTIR检测采用KBr压片法, KBr与样品质量比200:1, 扫描范围4 000~400 cm-1。 XPS使用带单色器的镁靶(Mg Kα , 光子能量1 253.6 eV), 分析室真空度5× 10-9 Pa, 测试所得的XPS峰以C(1s)(284.8 eV)为基准校正。 利用2D-Shige软件绘制2D-IR谱图, Origin软件处理其他数据并绘图。
不同胁迫强度下金盏菊原生质体Pb/Cd赋存形态分布见表1。 原生质体可交换态Pb/Cd所占比重一直不高, 比如在Pb/Cd胁迫强度为600/30 mg· kg-1时(处理D), 可交换态Pb/Cd分别占其全量的13.54%/29.59%。 随着Pb/Cd浓度的增大, 原生质体有机结合态Pb比重迅速升高, 而各形态Cd含量变化均不明显, 这可能与原生质体内转运体蛋白、 有机酸等组分对Pb/Cd的存贮和解毒机制有关。 此外, Dai[5]之前证实细胞壁对Cd的优先截留效应, 可能也在一定程度上调控了Cd/原生质体的结合。
| 表1 不同胁迫强度下金盏菊原生质体Pb/Cd赋存形态 Table 1 Speciation of lead/cadmium in Calendula Officinalis protoplast at different lead/cadmium concentrations |
不同胁迫强度对原生质体XRD图谱的影响如图1所示。 总体上看, 图1(a)中四组图谱差异不大, 具有某些共性特征。 其中, 最强信号均出现在31.7° 左右, 代表原生质体系NaCl晶体特征峰[6]; 26.7° 和45.5° 附近发现若干次峰, 分别鉴定为Pb盐 [Pb5(PO4)3Cl]和Cd盐晶体(CdS)特征峰[7]。 通常认为, 植物体内的盐分(NaCl等)具有维持自身生理代谢、 调控体系水势和渗透势平衡的作用; 植物可能通过摄取较多盐基离子存于体内, 以便缓解重金属毒害和胁迫效应[8]; 而进入原生质体的重金属则常以螯合固定、 胞内沉淀等方式降低活性和毒性。 将XRD图谱局部放大[见图1(b)], 发现胁迫后谱中29.4° 衍射峰几近消失; 这个特征峰代表了含Mg组分的存在, 推测金盏菊在吸收Pb/Cd的同时, 可能发生了离子交换或元素外排等生理响应。
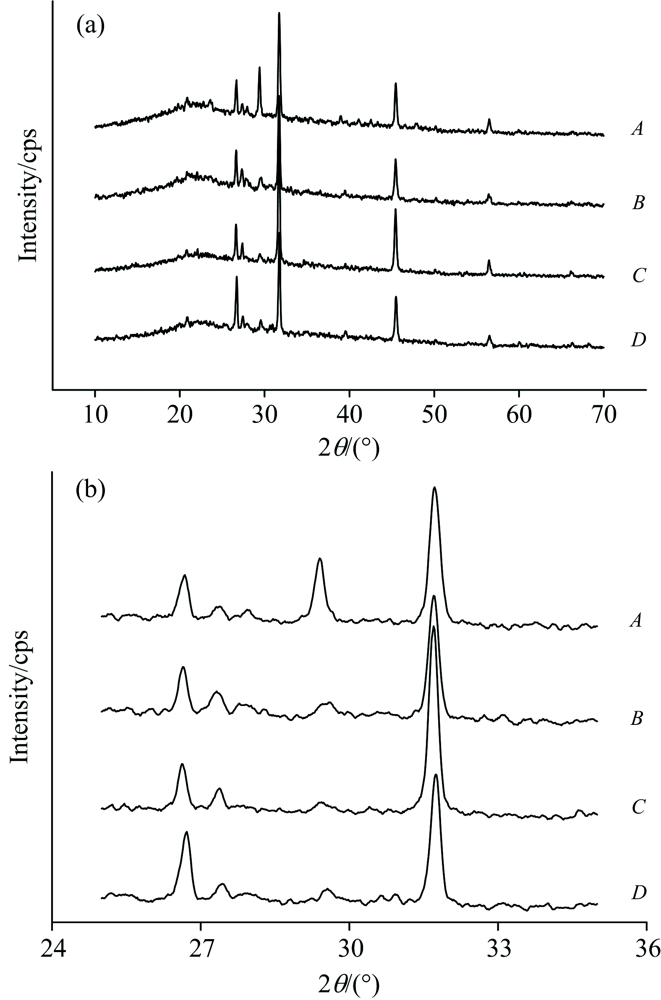 | 图1 不同胁迫强度下金盏菊原生质体X-衍射光谱Fig.1 XRD patterns of Calendula Officinalis protoplast at different lead/cadmium concentrations |
不同胁迫强度对原生质体FTIR图谱的影响见图2。 FTIR图谱总体走势比较相似, 但细节差异仍很明显。 其中: 位于3 510 cm-1附近的强吸收带代表— OH伸缩振动, 源于原生质体多糖、 蛋白质、 可溶性酶等组分; Pb/Cd胁迫使得该区域峰型复杂, 峰位不同程度偏移, 暗示了胁迫过程对金盏菊代谢的干扰效应, 以及可能导致的原生质体性质异化[9]。 1 650 cm-1附近为可溶性果胶— COOR和可溶性蛋白组分C=O (酰胺化合物Ⅰ 带和Ⅱ 带)特征峰, 胁迫过程对该处波峰影响很小, 说明金盏菊能够一定程度上适应重金属的逆境胁迫, 推测响应过程可能比较复杂。 事实上, 已有证据表明: 植物可通过蛋白组分(富甘氨酸蛋白、 富脯氨酸蛋白等)的诱导合成调控自身代谢并应对重金属胁迫[10]。 但胁迫程度过大(曲线D)时, 该处波峰发生变形或几近消失, 证明金盏菊对重金属的隐忍能力有限。 此外, 图谱中可辨识的重要波峰还包括: 脂族烃C— H弯曲振动峰(1 384 cm-1)、 类多糖组分C— O不对称伸缩振动(1 068 cm-1)等, 但胁迫过程对其峰位、 峰强干扰较小。 笔者之前掌握了金盏菊细胞壁对Pb/Cd胁迫的应答光谱[4], 经与本部分结果对比, 发现原生质体— OH和C=O可能对Pb/Cd胁迫的敏感性更强。
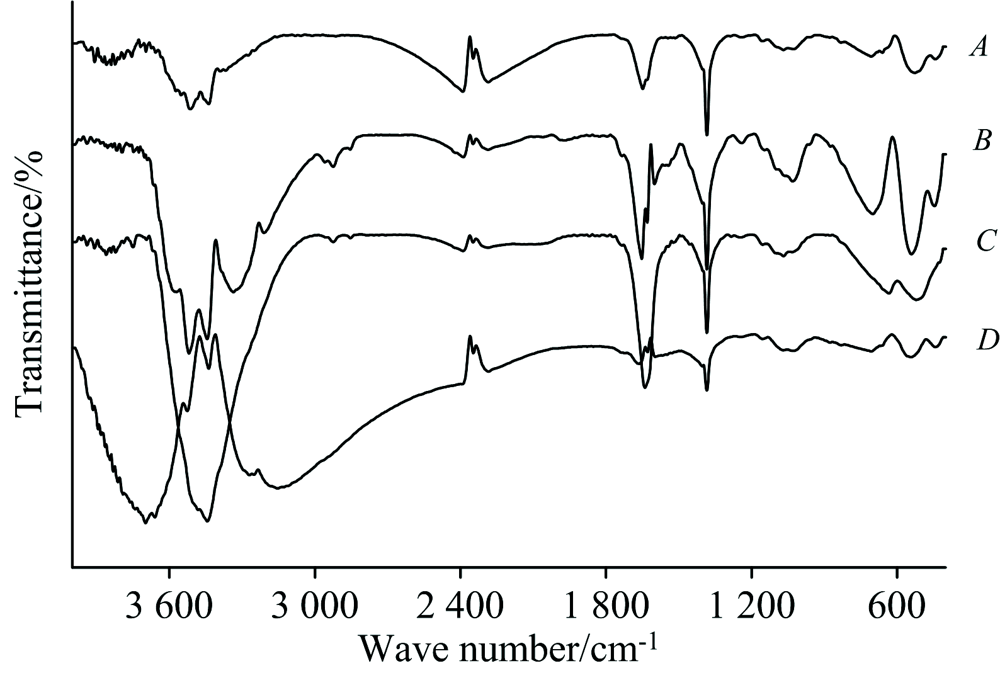 | 图2 不同胁迫强度下金盏菊原生质体红外光谱Fig.2 FTIR spectra of Calendula Officinalis protoplast at different lead/cadmium concentrations2.4 2D-IR |
2D-IR能够提高光谱分辨率, 并揭示官能团间的相互作用。 依据Noda规则[11]深入解析不同胁迫强度下金盏菊原生质体的2D-IR图谱, 结果见图3。 在同步图谱[图3(a)]中, 4 000~2 000 cm-1波段发现5个强自动峰, 分别为3 514 cm-1(— OH伸缩振动)、 3 438 cm-1(— CONH2)、 2 924 cm-1(脂族碳— CH3, — CH2伸缩振动)、 2 389 cm-1(氨基糖C≡ N伸缩振动)和2 285 cm-1(三键或累积双键)附近; 其中3 514 cm-1为最强峰, 说明— OH对Pb/Cd胁迫的反馈信号最敏感。 图3(b)的异步图谱中, 交叉峰(3 650/2 389), (3 438/2 924)和(2 924/2 389) cm-1符号为正, 表明在反应过程中, Pb/Cd优先与— OH结合, 次优先与脂族碳(— CH3, — CH2)结合, 与C≡ N的结合优先度最差; 交叉峰(3 438/2 389)符号为负, 说明3 438和2 389 cm-1峰强在Pb/Cd胁迫条件下变化方向相反(即一个峰强增加, 另一个峰强减小), 说明两种氢键存在相互转化的现象[12]。
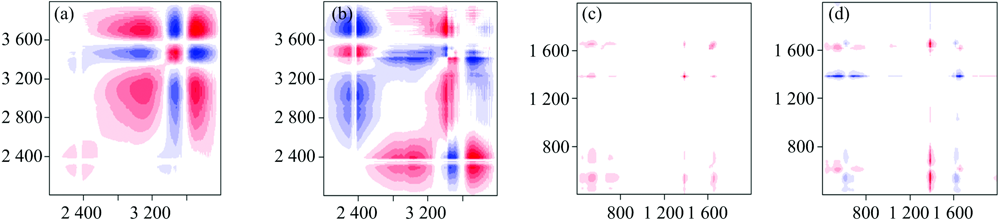 | 图3 不同胁迫强度下金盏菊原生质体二维相关红外光谱 (a), (c): 同步图谱; (b), (d): 异步图谱Fig.3 2D-IR correlation spectra of Calendula Officinalis protoplast at different lead/cadmium concentrations (a), (c): Synchronous map; (b), (d): Asynchronous map |
在2 000~400 cm-1的同步图谱[图3(c)]中, 获得的强自动峰顺序为1 383 cm-1(果胶质糖醛酸C— O)> 1 649 cm-1(酰胺化合物Ⅰ 带中C=O> 526 cm-1(C— H变形)> 708 cm-1(苯环C— H), 即Pb/Cd胁迫对C— O和C=O影响较大, 对苯环C— H影响最小(注: 符号“ > ” 表示前面的先于后面的吸收峰发生变化, 下同)。 在图3(d)的异步图谱中, 出现的交叉峰(1 383/526), (1 383/708), (1 649/526), (708/526)符号为正, 交叉峰(1 649/1 383), (1 649/526)符号为负, 说明官能团的总体变化顺序为1 649> 1 383> 526 cm-1。 综上可知, Pb/Cd胁迫对金盏菊原生质体官能团的影响优先作用于— OH和C=O, Pb/Cd将优先与这两类官能团反应结合。
不同胁迫强度对金盏菊原生质体XPS图谱的影响见图4。 对于A(CK)处理样品, XPS全谱的主要光电子峰分别代表C(1s)和O(1s), 结合能分别为284.77和532.41 eV。 结合相关文献[13], 认定C(1s)和O(1s)对应的官能团分别为C— OH和— COOH。 C(1s)和O(1s)都仅出现1个峰, 说明C原子和O原子基本处于同一化学环境。 胁迫反应后, 图谱光电子峰分别为C(1s), O(1s), Pb(4f)和Cd(3d)能级的四种元素。 为了深入揭示各元素结合能的变化, 对XPS分谱进行绘制。
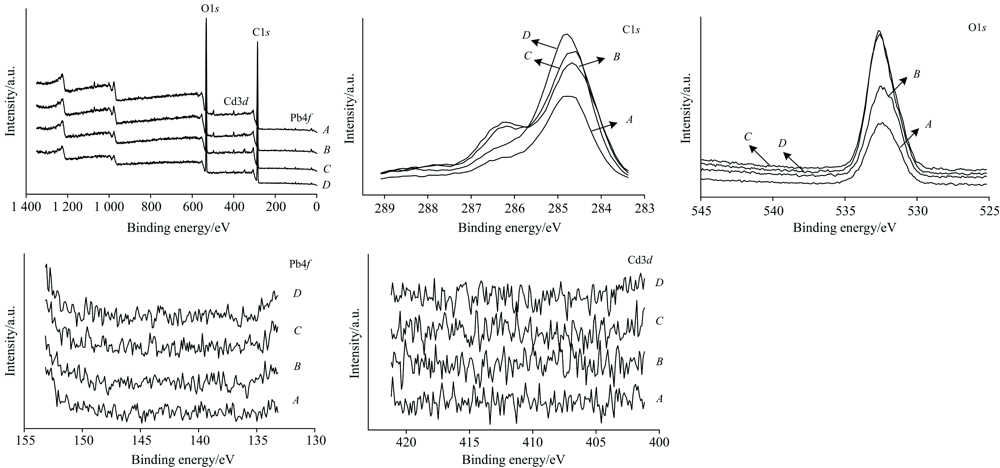 | 图4 不同胁迫强度下金盏菊原生质体X射线光电子能谱Fig.4 XPS spectra of Calendula Officinalis protoplast at different lead/cadmium concentrations |
在不同胁迫强度下, C(1s)结合能分别为284.79 eV(处理B)、 284.79 eV(处理C)和284.78 eV(处理D)。 Pb/Cd胁迫导致C(1s)结合能略有增加, 说明胁迫前后原生质体C原子的化学环境变化很小, C原子一定程度上参与了配位反应[14]。 O(1s)结合能分别为532.36 eV(处理B)、 532.70 eV(处理C)和532.68 eV(处理D), 峰位发生不同程度偏转。 对于处理B, O(1s)峰降低了0.05 eV, 源于O原子在反应过程获得电子, 使自身结合能降低, 这说明酚羟基、 羧基、 内酯基等含O基团参与了反应过程。 对于处理C和处理D, O(1s)的光电子峰分别向高场偏移了0.29和0.27 eV; 边界轨道理论和Pearon软硬酸碱理论认为: 光电子峰由低结合能向高结合能移动, 表明系统势能增大, 系统间发生了软-软作用和电子轨道交叠过程[15]。 据此可知, 本研究中含O官能团对Pb/Cd的结合过程可能涉及多种作用途径。
胁迫反应导致XPS图谱出现Pb(4f)和Cd(3d)光电子峰。 其中Pb(4f)峰定位于133.58 eV(处理B)、 137.78 eV(处理C)和141.38 eV(处理D)附近, 这是π 电子-Pb离子相互作用的结果[16]。 Cd(3d)峰分布于402.58 eV(处理B)、 410.68 eV(处理C)和417.68 eV(处理D)附近; 随着胁迫浓度的增加, Cd(3d)结合能逐渐增加, 暗示了反应过程中Cd离子具有明显失电子倾向, 是失电子一方。 图谱中Pb(4f)和Cd(3d)光电子峰识别效果不明显, 可能源于响应信号较弱; 经调整测试条件重复实验后, 基本排除了元素分布不均的情况, 但图谱改进效果仍然很小。 据此认为, 导致信号较差的原因在于原生质体Pb/Cd含量较低。
在长期的选择进化过程中, 植物自身对重金属胁迫产生了天然抗性和耐受性。 植物可以调控地下根部的生理生化行为(根际环境、 根系分泌物、 重金属形态等)来阻断污染物的运输路径, 也可以通过更为复杂的传导信号(转运蛋白、 蒸腾作用、 液泡解毒等)滞缓污染物的地上部长距离运输。 本研究不以分子生物学为切入点, 而从光谱层面入手, 对重金属与植物原生质体间的反应机制进行分析, 同样得到了有说服力的一手数据。 可以看出, 金盏菊原生质体与Pb/Cd的响应行为相对复杂: 一方面, 原生质体中检测出Pb/Cd的存在, 且其赋存形态随着胁迫强度的差异出现不同程度的分化; 另一方面, Pb/Cd在进入原生质体后, 或以金属沉积物(Pb盐、 Cd盐等)形式存留, 或与官能团(— OH、 C=O等)相互作用共存, 或通过元素外排(比如Mg元素)产生信号响应。 这与同类研究有吻合之处, 也弥补了谱学解析层面的短板和欠缺。 本结果能与分子生物学证据互为补充, 协同验证重金属/植物原生质体的反馈调节机制, 但以细胞膜、 液泡为代表的细胞器等精细结构对重金属胁迫的抵抗贡献仍需深度发掘。 后续研究将据此展开。
金盏菊原生质体对Pb/Cd胁迫响应的谱学信号比较显著, 胁迫强度与Pb/Cd赋存形态具有不同程度的内在关联。 XRD图谱最强峰位于31.7° (NaCl晶体), Pb盐[Pb5(PO4)3Cl]和Cd盐(CdS)特征峰也很明显。 FTIR和2D-IR结果显示: 3 510 cm-1附近强吸收带为— OH伸缩振动峰, Pb/Cd胁迫导致峰形复杂、 峰位偏移, — OH和C=O表现出与Pb/Cd的优先结合趋势。 Pb/Cd胁迫导致XPS图谱出现Pb(4f)和Cd(3d)特征峰, 其中Pb(4f)峰源于π 电子-Pb的结合效应, Cd(3d)结合能升高说明Cd具有失电子倾向。 光谱识别技术可以有效判别原生质体与Pb/Cd接触的反馈信号, 对于挖掘植物体内的重金属运输途径、 耐受响应和生理解毒机制, 构建原生质体融合高效菌群以及剖析重金属/原生质体生态毒理学具有重要价值。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|