{kind=link}
{kind=link}
{kind=link}
{kind=link}
钾镁胁迫对橡胶幼苗叶片物质成分影响的FTIR表征
[薛欣欣 , 王文斌, 罗雪华, 张永发, 赵春梅]
, 王文斌, 罗雪华, 张永发, 赵春梅]
, 王文斌, 罗雪华, 张永发, 赵春梅]
|
|


作者简介: 薛欣欣, 1986年生, 中国热带农业科学院橡胶研究所助理研究员 e-mail: xuexinxin_2010@163.cm
养分亏缺是作物产量和质量提升的重要限制因子。 我国砖红壤植胶区钾、 镁缺乏及其低的有效性长期制约天然橡胶的产、 质量。 以“热研7-33-97”橡胶幼苗为研究对象, 利用傅里叶变换红外光谱(FTIR)对钾、 镁胁迫下橡胶树叶片物质组成特征进行研究, 分析不同钾、 镁胁迫条件下叶片红外光谱图谱特征峰的变化及其差异, 探讨钾、 镁胁迫对叶片物质成分和结构的影响, 以期为钾镁缺乏影响橡胶树生长与生理代谢的机理研究提供参考。 结果表明: (1)缺钾条件下, 1 554, 1 519和1 075 cm-1处的吸收峰缺失, 表明橡胶叶片中蛋白质酰胺Ⅱ带、 酚类物质、 碳水化合物的化学结构受到破坏, 并且其他特征峰吸光度与正常处理相比均有不同程度的升高, 表明缺钾造成蛋白质、 碳水化合物等物质在叶片中大量积累, 物质转运效率降低; (2)缺镁条件下, 1 554 cm-1处吸收峰向高频方向位移了6 cm-1, 说明缺镁造成蛋白质酰胺Ⅱ带的结构发生改变, 同时, 1 550~1 350 cm-1波数范围内吸收峰相对吸光度明显下降, 说明缺镁降低了细胞壁多糖以及含油脂化合物的含量, 且该波数范围内的三个吸收峰较其他吸收峰变化敏感, 表明该波段可以较好地指示橡胶叶片镁营养状况; (3)钾镁同时缺乏条件下, 各吸收峰的相对吸光度和强度明显减弱, 1 554, 1 519和1 075 cm-1处的吸收峰缺失, 说明蛋白质酰胺Ⅱ带、 酚类物质、 碳水化合物的化学结构受到破坏, 1 057 cm-1处吸收峰向高频方向位移了11 cm-1, 表明橡胶叶片中果胶类多糖分子结构发生了变化。 综上所述, 缺钾造成橡胶叶片蛋白质、 糖类物质的含量的大量积累, 缺镁造成叶片细胞壁多糖和油脂化合物下降, 而钾镁同时缺乏时蛋白质、 脂类、 糖类物质等物质含量均明显下降。 研究表明, 使用FTIR技术对养分缺乏下的橡胶叶片物质成分定性分析具有一定的可行性, 同时可为橡胶钾、 镁营养生理代谢的机制研究提供新的思路和方法。
Nutrient deficiency is a vital limiting factor for crop yield and quality improvement. It has restricted the yield and quality of natural rubber for a long time because of the depletion of soil potassium (K) and magnesium (Mg) and their low plant availability in the Laterite soil of rubber plantations, South China. Rubber seedlings of “CATAS 7-33-97” was chosen as test material, the material composition characteristics of rubber leaves under K and Mg deficiency were studied by Fourier transform infrared spectroscopy (FTIR). The changes and differences of FTIR characteristic peaks of leaves were analyzed, and the material compositions and structures were discussed. It would provide some physiological evidence of rubber plants subjected to K and Mg deficiency stress. The results showed that: (1) the characteristic peaks of 1 554, 1 519 and 1 075 cm-1 disappeared due to K deficiency, implying that the structure of amide Ⅱ band, phenolic compounds and carbohydrate were broken. The relative absorbance of all characteristic peaks was increased compared with normal treatments, suggesting that protein and carbohydrate were accumulated in leaf to reduce their translocation efficiency. (2) The absorption peak at 1 554 cm-1 shifted 6 cm-1 to high frequency as 1 560 cm-1 when the plant was subject to Mg deficiency, which indicated that the structure of the protein amide Ⅱ band was changed. Meanwhile, Mg deficiency decreased the relative absorbance in the range of 1 550~1 350 cm-1, implying that Mg deficiency decreased the contents of polysaccharides and lipid compounds of the leaf cell wall. The three absorption peaks in the wave number of 1 550~1 350 cm-1 were more sensitive to Mg deficiency than other absorption peaks, indicating that the wave band could better represent the magnesium nutrition status of rubber plant leaves. (3) The relative absorbance and intensity of each absorption peak decreased obviously at both K and Mg deficiency. Absorption peaks at 1 554, 1 519 and 1 075 cm-1 disappeared similar to that K deficiency, which indicated that the chemical structure of protein amide Ⅱ band, phenols and carbohydrates was destroyed. Meanwhile, the absorption peak of 1 057 cm-1shifted to the high frequency direction by 11 cm-1 to 1 068 cm-1, implying the molecular structure of pectin polysaccharides changed in rubber leaves. In conclusion, K deficiency caused a large accumulation of protein and carbohydrates in rubber leaves, and magnesium deficiency caused the decrease of polysaccharide and lipid compounds in the leaf cell wall. However, the contents of protein, lipid and carbohydrates decreased significantly when the plant was subjected to both K and Mg deficiency. The results showed that FTIR technology was feasible for qualitative analysis of the material components of rubber leaves under nutrient deficiency and could provide new ideas and methods for studying the mechanism of K and Mg nutrition and physiological metabolism of the rubber trees.
天然橡胶是重要的战略物资和工业原料, 在国民经济发展中具有非常重要的地位和作用。 橡胶树(Hevea brasiliensis)是天然橡胶的重要来源, 为我国热带和亚热带地区重要的经济作物。 然而, 我国橡胶林的大规模垦植、 刺激割胶应用、 化肥施用不当等造成胶园土壤肥力出现下降趋势[1, 2]。 研究表明, 海南植胶后土壤速效钾含量仅为未开垦土壤的50%[3]。 橡胶缺钾会造成碳水化合物转运受阻、 叶片结构(面积、 厚度、 叶绿体结构等)发生变化[4], 进而影响橡胶树产、 排胶及橡胶质量[5]。 海南作为我国最大的热区之一, 其砖红壤具有阳离子交换能力低、 强风化、 酸性和砂质等特点, 土壤中水合半径较大的镁离子存在较高的淋失风险[6]。 研究发现, 海南植胶区80%以上的土壤交换性镁含量低于正常值[7]。 橡胶缺镁会造成暴光老叶叶脉间呈鱼骨褪绿而变黄, 严重时出现黄叶落叶病, 影响产量形成[8]。
傅里叶变换红外光谱(Fourier transform infrared spectroscopy, FTIR)技术是一种基于化合物中官能团和极性键振动的结构分析技术, 可对样品进行定性和定量分析, 具有灵敏度高, 制样方法简单, 样品用量少, 测试时间短等优点[9]。 植物体内生物大分子如核酸、 脂类和碳水化合物, 具有特征性的官能团和独特的分子振动模式, 通过分析红外光谱吸收位置、 宽度和强度, 有助于了解植物体内有机物分子官能团的组成和结构特征。 以往研究认为, 植株组织的FTIR吸收峰特征会因营养状态的不同而产生差异[10, 11]。 Butler等[12]证实FTIR能够对缺钙胁迫下模式植物鸭跖草的营养状态进行精确评估。 王盛峰等[13]利用FTIR研究了缺锌条件下玉米碳水化合物、 脂类、 蛋白质及核酸等组分发生了改变。 有报道利用FTIR研究了低硼和高硼胁迫下影响柑橘叶片物质组成和结构。 有工作对棉花功能叶片FTIR谱图特征峰的研究发现缺钾破坏了叶片表皮结构, 蛋白质、 纤维素、 可溶性糖和核糖等结构发生变化, 缺硼造成蛋白质及可溶性糖、 纤维素等碳水化合物在叶片大量积累。 姚宇洁等[14]利用FTIR研究了枳橙实生苗缺铁胁迫下叶片蛋白质、 纤维素、 可溶性糖和核糖结构发生了变化而且含量降低。 尽管如此, 利用FTIR技术对橡胶叶片物质成分构成及其对钾、 镁胁迫的响应鲜有报道。 本研究以橡胶树幼苗作为试验材料, 采用FTIR技术来表征钾镁胁迫下橡胶叶片化学组分的变化特征, 以期为橡胶钾、 镁生理代谢的机制研究提供新的思路和方法。
供试材料为生长一致且状态良好的橡胶(Hevea brasiliensis)幼苗, 品系为热研7-33-97。 试验于2017年10月—2018年1月在中国热带农业科学院橡胶研究所进行。 将冲洗干净的幼苗固定在双孔黑色泡沫板上, 再移栽到盛有5 L营养液的黑色塑料桶中, 使泡沫板漂浮在营养液表面, 同时保证幼苗根部完全浸没在营养液中, 每盆定植2株, 置于人工气候箱中进行培养。 人工气候箱内设常温(25 ℃/20 ℃)(昼/夜), 光照强度(5 000 lx), 光周期为12 h/12 h(昼/夜), 相对湿度85%。 营养液配方参考Hogland and Aron(略有调整)。 大量元素(mg· L-1): KCl, 596; MgSO4, 240; Ca(NO3)2, 656; (NH4)2SO4, 462; NaH2PO4, 120; 微量元素(mg· L-1): MnSO4, 15; H3BO3, 6.2; ZnSO4, 5.0; Na2MoO4, 0.21; CuSO4, 0.016; EDTA-Fe, 50 μmol· L-1。 设正常(全营养液CK)、 缺钾(-K)、 缺镁(-Mg)和钾镁同时缺乏(-K-Mg)四个处理, 3次重复, 每重复10盆、 共20株幼苗, 为维持营养液中离子浓度平衡, 缺素处理采用NaCl和(Na)2SO4来调整Cl-和
叶片样品采集和处理: 橡胶幼苗培养60 d后, 选取橡胶幼苗顶端完全展开叶片, 一级水冲洗干净后将样品置于105 ℃烘箱中杀青30 min, 之后在75℃烘箱中烘干至恒重, 磨细并过100目筛, 备用。
样品测定: 采用溴化钾压片法。 压片之前, 将溴化钾(光谱纯)粉末放入烘箱烘干至恒重, 然后分别称取叶片粉末样品和溴化钾(光谱纯)0.200和2.000 g(质量比1:10)加到玛瑙研钵中, 在红外灯照射干燥条件下按同一方向磨细呈淀粉状, 取少许样品, 在真空条件下, 用压杆缓慢加压至约15 MPa, 维持1 min左右, 得到样品薄片; 采用上述方法, 将溴化钾制成薄片作为背景。 将薄片放在Nicolet公司的Inpact-410型FTIR光谱仪上进行测定, 测定波数范围4 000~400 cm-1, 扫描累加次数32, 分辨率4 cm-1, 每个样品测定前均对背景进行扫描, 去除背景的影响。
采用OMNIC8.2软件对叶片样品的FTIR谱图进行数据处理, Origin8.0软件作图。
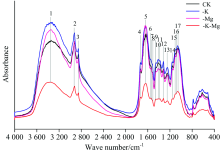
橡胶幼苗叶片的红外光谱图如图1所示。 各化学键的吸收处于中红外区, 即波数为4 000~400 cm-1。 各处理下的叶片具有一些典型的共有吸收峰, 但各处理间的特征峰数目以及相对吸光度存在不同程度差异。 结合表1进一步分析发现, 与正常处理(CK)相比, 缺钾(-K)条件下, 橡胶叶片1 554, 1 519和1 075 cm-1处的吸收峰缺失, 并且3 345和1 541 cm-1处的吸收峰分别向高频方向位移了9和6 cm-1后分别位于3 354和1 547 cm-1处; 缺镁(-Mg)条件下, 橡胶叶片1 075 cm-1处的吸收峰缺失, 并且3 345, 1 631, 1 554和1 057 cm-1处的吸收峰分别向高频方向位移了8, 4, 6和11 cm-1; 钾镁同时缺乏(-K-Mg)条件下, 1 554, 1 519和1 075 cm-1处吸收峰缺失, 并且3 345和1 541 cm-1处的吸收峰向高频方向位移了8和5 cm-1。 由此可见, 钾、 镁营养缺乏下, 橡胶叶片物质组分结构发生了变化。
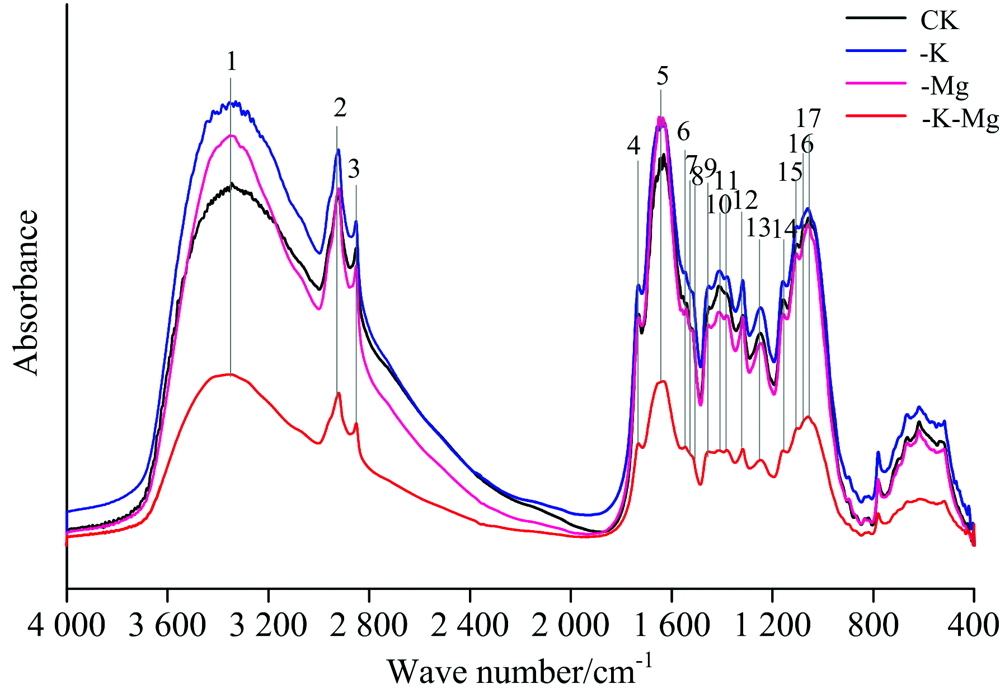 | 图1 不同处理下橡胶叶片的FTIR全谱图(4 000~400 cm-1)Fig.1 FTIR spectra of the leaves of rubber seedlings under different treatments (4 000~400 cm-1) |
| 表1 不同处理下橡胶叶片的特征峰 Table 1 Characteristic peak of FTIR in rubber leaf under different treatment |
4 000~400 cm-1范围, 钾镁同时缺乏条件下特征峰的相对吸光度明显低于其他处理, 且特征峰吸收强度明显减弱; 缺钾条件下的特征峰相对吸光度高于正常处理; 在4 000~1 600 cm-1波数范围, 缺镁条件下的特征峰相对吸光度高于正常处理, 而波数处于1 600~400 cm-1时, 缺镁则低于正常处理。 由此表明, 钾、 镁营养缺乏时, 橡胶叶片各物质组分的含量发生了改变。 由于1 800~900 cm-1波数范围内特征峰的数量较为集中, 且相对吸光度变化幅度最显著, 为提高谱图的分辨率以减少叠加, 对不同处理下1 800~900 cm-1波数范围内的FTIR谱图做进一步分析。
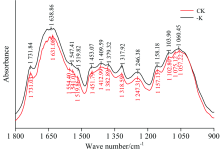
由图2可知, 与正常处理(CK)相比, 缺钾(-K)条件下橡胶叶片在1 800~900 cm-1范围内的吸收峰数量、 吸光度以及吸光强度等方面均发生了明显的变化, 其中, 1 554和1 075 cm-1处的特征峰缺失, 结合红外光谱特征峰归属表(表2)可知, 1 554 cm-1附近表征总蛋白酰胺Ⅱ 带吸收, 为C—N键和N—H键的弯曲振动, 1 075 cm-1附近表征碳水化合物C—O键的伸缩振动, 由此可见, 低钾胁迫破坏了蛋白质及碳水化合物的分子结构, 这与郝艳淑在棉花上的研究结果相似[15]。 此外, 相比正常处理下的1 541.01 cm-1的吸收峰, 缺钾胁迫造成该吸收峰向高频方向移动6.40 cm-1, 1 541 cm-1为蛋白质酰胺Ⅱ 带, 说明缺钾胁迫影响了橡胶叶片中C—N键和N—H键的结合, 造成蛋白质的结构发生改变。
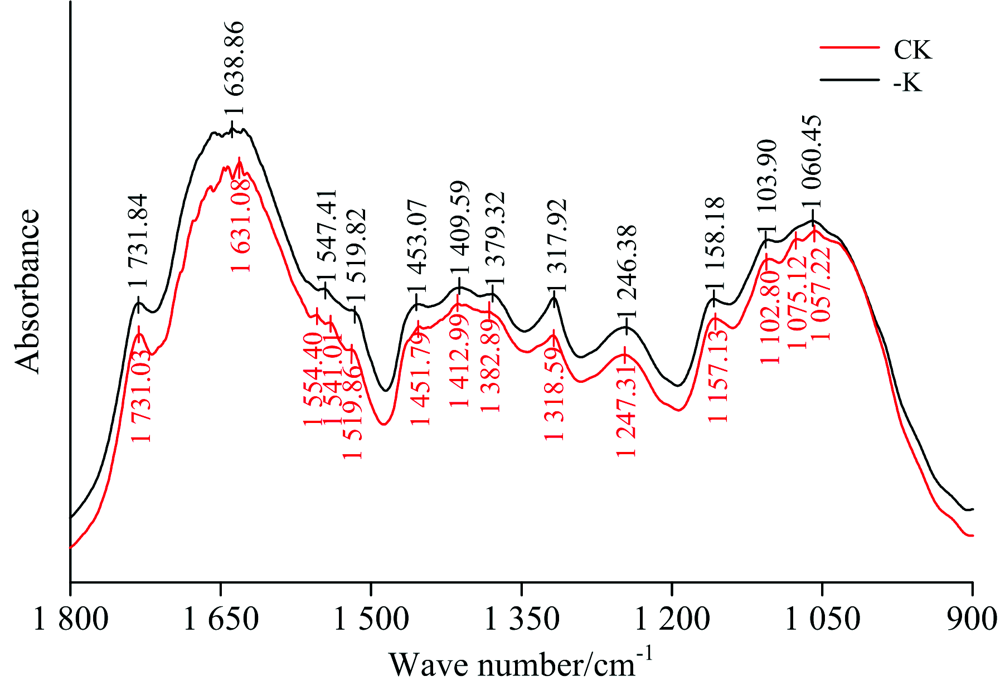 | 图2 缺钾条件下橡胶叶片的FTIR光谱特征(1 800~900 cm-1)Fig.2 FTIR spectra of the leaves of rubber seedlings under K deficiency (1 800~900 cm-1) |
| 表2 叶片官能团和相应的红外吸收频率 Table 2 Functional groups of the leaf and the corresponding infrared absorption frequencies |
由图2可明显看到, 缺钾胁迫时各特征峰的相对吸光度均高于正常处理, 说明缺钾胁迫影响叶片蛋白质及可溶性糖、 纤维素、 果胶、 酯类等碳水化合物的积累, 并造成光合产物向韧皮部的运输或转载受阻, 导致这些大分子物质较多的积累在叶片中, 进而影响植株的正常生长发育。 该结果与相关研究相似, 但与有些研究结果相反。 研究认为, 缺钾条件下棉花功能叶片蛋白质和碳水化合物纤的含量均降低。 分析以上结果差异的可能原因, 一方面与缺钾造成光合作用减弱, 蛋白质和碳水化合物合成受阻有关[16], 另一方面也可能与品种的钾效率高低有关(钾高效品种降低、 低效品种积累)[15]。 以往研究通过化学分析方法也证实了本研究结果, 即缺钾橡胶叶片中可溶性糖含量显著增加、 叶绿体中的淀粉粒明显增多, 导致碳水化合物在叶片中积累, 阻碍了其向其他器官的运输[4]。
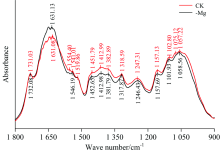
由图3可以看出, 与正常处理(CK)相比, 缺镁(-Mg)胁迫下1 075 cm-1处的吸收峰缺失, 结合红外光谱特征峰归属(表2)可知, 1 075 cm-1附近的吸收峰表征碳水化合物的C—O伸缩振动吸收, 表明缺镁胁迫造成C—O键受到破坏, 进而改变了碳水化合物的分子结构。 另外, 与正常处理相比, 缺镁胁迫增加了1 631 cm-1峰的吸收强度, 1 631 cm-1表征酰胺Ⅰ 带的C=O伸缩振动, 说明, 缺镁胁迫增加了橡胶叶片蛋白质酰胺Ⅰ 带的含量, 但降低了1 554 cm-1的吸光强度, 1 554 cm-1附近代表蛋白质的酰胺II带的C—N伸缩振动和N—H弯曲振动。 此外, 1 554 cm-1处吸收峰向高频方向位移了6 cm-1, 说明缺镁胁迫造成蛋白质酰胺Ⅱ 带的结构发生了改变。
 | 图3 缺镁条件下橡胶叶片的FTIR光谱特征(1 800~900 cm-1)Fig.3 FTIR spectra of the leaves of rubber seedlings under Mg deficiency (1 800~900 cm-1) |
由表2可知, 1 731, 1 519和1 318 cm-1处吸收峰分别代表细胞壁果胶成分中脂类化合物的C=O伸缩振动吸收、 酚类物质的苯环骨架振动吸收以及纤维素的CH2面外摇摆振动吸收。 与正常处理相比, 缺镁胁迫对以上吸收峰的吸光度和位置均无明显影响, 由此说明缺镁胁迫并不影响细胞壁果胶中的脂类化合物、 酚类物质以及纤维素含量和结构。 从1 550~1 350 cm-1的特征峰吸光度变化分析, 缺镁胁迫处理明显低于正常处理, 且相比其他吸收峰的降幅较大, 说明以上波段吸收峰变化对缺镁胁迫较为敏感, 可以作为橡胶镁营养变化的特征光谱识别区域; 该区间内的三个特征吸收峰分别代表细胞壁多糖的C—H弯曲振动、 酯类化合物的—COO伸缩振动和纤维素类物质的C—H弯曲振动及CH3的剪式振动, 由此也说明缺镁胁迫明显减少了橡胶叶片细胞壁多糖以及含油脂化合物等物质含量。 以往有关研究也证实了利用红外光谱技术快速无损诊断黄瓜镁元素亏缺是可行的[17], 但在橡胶上的应用鲜有报道, 仍需更深入开展研究。
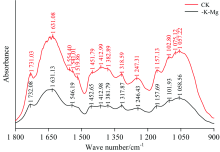
分析图4, 与正常处理(CK)相比, 钾镁同时缺乏(-K-Mg)条件下橡胶叶片FTIR图谱的吸收峰明显变少, 并且吸收峰的吸光强度明显减弱, 由此说明钾镁同时缺乏造成橡胶叶片中蛋白质、 脂类、 纤维素、 可溶性糖等含量明显下降以及某些物质的结构发生改变。 进一步分析发现, 1 554, 1 519和1 075 cm-1吸收峰缺失, 这与缺钾胁迫较为相似, 其中1 554 cm-1吸收峰附近为蛋白质酰胺Ⅱ 带的C—N伸缩振动, 1 519 cm-1为酚类物质的苯环骨架振动, 1 075 cm-1为碳水化合物的C—O伸缩振动, 说明钾镁同时缺乏破坏了蛋白质酰胺Ⅱ 带、 酚类物质以及碳水化合物的化学结构。 另外, 钾镁同时缺乏造成1 541.01 cm-1向1 546.19 cm-1位移了5.18 cm-1, 1 541 cm-1附近的吸收峰为蛋白质酰胺Ⅱ 带的N—H弯曲振动, 进一步也说明钾镁同时缺乏造成蛋白质酰胺Ⅱ 带的结构发生改变。
 | 图4 钾镁同时缺乏条件下橡胶叶片的FTIR光谱特征(1 800~900 cm-1)Fig.4 FTIR spectra of the leaves of rubber seedlings under both K and Mg deficiency (1 800~900 cm-1) |
采用傅里叶红外光谱测定了不同钾、 镁胁迫处理下的橡胶叶片化学物质组成。 得到以下结论: (1)缺钾造成橡胶叶片中蛋白质酰胺Ⅱ 带、 酚类物质、 碳水化合物的化学结构受到破坏, 以及蛋白质、 碳水化合物等物质在叶片中大量积累; (2)缺镁降低了细胞壁多糖以及含油脂化合物的含量, 1 550~1 350 cm-1波段内的吸收峰可以较好地指示橡胶叶片镁营养状况; (3)钾镁同时亏缺造成橡胶叶片中果胶类多糖分子结构发生了变化, 蛋白质、 脂类、 糖类物质等物质含量均明显下降。 本研究表明, 使用FTIR技术对养分缺乏下的橡胶叶片物质成分定性分析具有一定的可行性, 同时可为橡胶钾、 镁营养生理代谢的机制研究提供新的思路和方法。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|