
{kind=link}
{kind=link}
应用高光谱数据估算植物物种beta多样性
[彭羽1, 2, *  , 陶子叶
, 陶子叶2 , 许子妍2 , 白岚2 ]
, 陶子叶|
|


作者简介: 彭 羽, 1973年生, 中央民族大学生命与环境科学学院副教授
近年来, 光谱分析应用于植物多样性的估算引起了全球生物多样性学界的广泛关注。 基于光谱异质性假说(SVH), 大量案例研究应用光谱指数估算了森林、 草原等的植物物种alpha多样性, 但是beta多样性的研究尚缺乏。 在我国浑善达克沙地中部调查270个直径为0.8 m的植物群落样方, 测量植物物种beta多样性, 并采集样方高光谱数据(375~1 025 nm)。 中随机抽取样方数据165个作为模型训练数据, 105个作为模型验证数据。 beta多样性指数选用Bray-Curtis index (BC), Sörensen index (S) 和Jaccard index (J)。 基于物种特征波段, 开发了164个高光谱指数估算物种beta多样性指数。 采用Pearson相关性分析对开发的高光谱指数进行初步筛选, 然后比较不同植物群落盖度和群落复杂性条件下高光谱指数的稳定性, 进一步筛选。 结果表明, 400~1 000 nm光谱反射率一阶导数的相似性指数和欧氏距离指数, 以及760~800 nm之间的相似性指数, 能够较好地估算植物物种beta多样性。 其中, 物种BC指数与高光谱欧氏距离指数表现最为一致, 二者都考虑了物种组成数量的差异, 物种S和J指数拟合效果较差。 本研究对于促进高光谱应用于植物物种多样性估算具有推进作用。
Spectral analysis has been increasingly applied to estimate plant species diversity through the world, especially for biodiversity field. Although spectral variability hypothesis (SVH) has been widely proved in estimating plant alpha diversity for tropical, temperate, and sub-tropical forests, meadow, steppe and grasslands, however, the performance on beta diversity is still lack. In this study, we measured the hyperspectral reflectances and plant species diversity indices of 270 plots at a fine scale (0.8 meter) in central Hunshandak sandy grasslands of Inner Mongolia, China. 195 plots were used as training data and 75 plots as validating data. Bray-Curtis dissimilarity index (BC), Sörensen index (S) and Jaccard index (J) were calculated to indicate actual beta diversity. Based on spectral biological features of different plant species, 164 hyperspectral indices were developed and used to assess plant species beta diversity. Pearson’s correlation analysis and multiple linear stepwise regression were conducted based on sensitive wavebands to produce hyperspectral models. The hyperspectral indices which high Pearson’s correlation coefficients will be remained for further tested. Communities with different coverages and richness were also used to test the robustness of proposed models. By comparing the stability of hyperspectral indices under different communities, the indices with high stability is remained for validation by 75 plots. Results demonstrated that BC, Euclidean distances of first-order derivation values between 400~1 000 nm, and BC of 760~800 nm could accurately estimate species beta diversity. BC can be accurately estimated by hyperspectral indices, since they were both calculated as parameters of the distance between plots. The Jaccard and Sörensen indices were hardly estimated, it is hard to find the suitable wavebands or other parameters in spectral data to replace the “common reflectance” between pairwise plots. This study promotes the development of methods in assessing plant species beta-diversity using hyperspectral data.
生物多样性对于维持地球生态系统的稳定和保持生态系统服务功能具有重要意义[1]。 由于全球气候变化、 土地利用变化、 人为干扰增强和生物入侵等原因, 全球生物多样性正在下降, 预测未来也会进一步下降[2, 3]。 生物多样性的快速评估对于生物多样性监测和保持至关重要。 近年来, 光谱数据开始应用于植物物种多样性的快速评估[4]。 主要通过两种途径: 一是从光谱数据特征参数与物种多样性的关系进行直接评估, 二是从光谱数据衍生的环境变量、 实地调研数据和生物学特性对物种多样性进行间接评估[5, 6, 7, 8]。
光谱异质性假说(spectral variability hypothesis, SVH)认为, 由于不同物种的特征光谱参数不同, 植物物种多样性与光谱异质性直接相关[7, 9]。 基于SVH, 提取自不同生态系统类型, 包括温带森林、 热带雨林、 稀树草原、 山地草原、 干草原、 高草草原和沙地草原的光谱指数被应用于评估物种丰富度、 Shannon-weaver多样性指数和Simpson 多样性指数等alpha群落内物种多样性指数[10]。 Beta群落间或环境梯度物种多样性是生物多样性的另一个重要组分, 表示群落物种组成的差异, 用于指示物种的空间异质性和替代性。 与仅仅测量alpha多样性指数相比, beta多样性指数能够反映物种多样性空间分布的动态特征, 对于揭示植物群落的维持机制具有重要意义, 近年来为国内外学者所关注。 航空以及卫星遥感数据已经用于评估beta多样性, 但是由于光谱分辨率的限制, 此法还存在很大的不确定性。 高光谱数据具有光谱分辨率高、 光谱信息丰富的特点, 能够反映物种之间的差异, 已成功用于alpha多样性的评估[7, 8, 9, 10], 具有估算beta多样性的潜力。 然而, 目前还很少有高光谱估算beta多样性的案例研究。 我国北方的浑善达克沙地具有丰富的微尺度物种多样性, 适合开展植物物种beta多样性研究。 本研究在浑善达克沙地, 采集植物高光谱数据和实地样方调查数据, 计算beta多样性指数, 采用高光谱植被指数估算beta多样性。 本研究科学假设是植物特征波段光谱距离能够反映物种组成的差异。 基于收集到的270个不同盖度样方(< 10%, 11%~20%, 21%~30%, 31%~50%, > 50%)的植物物种数据, 探索以下问题: (1)能够估算beta多样性指数的最佳高光谱植被指数; (2)植物beta多样性指数与高光谱植被指数之间的关系是否受群落盖度和群落复杂性等条件的影响。
研究区位于浑善达克沙地中部的内蒙古自治区正蓝旗(41° 46'— 43° 69'N; 114° 55'— 116° 38'E)。 正蓝旗包含3个镇、 8个苏木和3个国营牧场, 2016年草场面积占72.3%。 该区属中温带半干旱大陆性季风气候, 年均温为1.7 ℃, 7月均温为18.7 ℃, 1月均温为-18.3 ℃, 年均降水为365 mm, 其中80%~90%的降水位于7月— 9月间。 该区主要由固定沙丘、 半固定沙丘、 流动沙丘和丘间低地、 湿地等景观组成。 植物物种多以沙生草本植物为主, 优势植物有沙蒿(Artemisia desertorum)、 冷蒿(Artemisia frigida)、 冰草(Agropyron cristatum)、 披碱草(Elymus dahuricus)等, 具有丰富的微尺度植物物种, 对于防治沙漠扩展具有重要作用。 该区丰富的生境、 相似的海拔和测量天气条件, 适合进行植物物种beta多样性高光谱估测的研究。
本研究在2017年7月到8月在浑善达克沙地中部内蒙古正蓝旗进行, 在固定沙丘、 半固定沙丘、 流动沙丘及低地4种生境的代表性区域共调查270个直径为0.8 m的圆形草本样方, 每个样方进行GPS定位, 统计每一样方内草本植物的种类、 数量、 高度和盖度等数据, 计算植物多样性指数。 采用三个beta多样性指数: Bray-Curtis dissimilarity index (BC), Sö rensen index (S)和Jaccard index (J)。 计算公式如式(1)
式(1)中, BC为样方间Bray-Curtis距离, xis和xjs分别表示植物物种s在样方i和j的多度, BC值介于0~1之间, 0表示两个样方物种组成完全一致, 1表示两个样方没有相同的物种。
S和J指数表示如式(2)和式(3)
式中, a和b分别表示两个样方中仅在一个样方出现的物种, c表示在两个样方均出现的物种。 S和J值介于0~1之间, 0表示两个样方物种组成完全不同, 1表示两个样方完全相同。 为了表示两个样方间的beta多样性, 采用1-S和1-J数值。
采用手持式地物光谱仪FieldSpec H2 (ASD Corp, USA)采集各个样方的冠层光谱。 仪器光谱范围为325~1 075 nm, 光谱分辨率为3 nm, 采样间隔为1 nm (www.asdi.com)。 光谱测量在风力小于3级的晴朗天气下进行, 采集时间为北京时间10:00— 15:00。 测量人员着黑色服装, 减少光谱干扰, 避免遮挡阳光。 光谱传感器探头垂直向下, 保持在冠层上方1.8 m内, 确保冠层充满仪器的视场角。 每一样本重复测量10次取均值。 测量前均同步测量参考白板反射的辐射光谱用于标定, 并定时进行系统优化, 以消除环境变化所带来的影响。 将获得的高光谱数据经滤波和去除噪音后, 计算一阶微分反射比(FD)。 FD分析用以消除土壤等环境背景值的影响, 提取不同植物群落的光谱特征, 用于拟合植物物种beta多样性指数。
基于SVH假说, 两个样方间的高光谱差异能够估算beta多样性。 光谱距离越近, 则光谱特征相似, beta多样性降低[7]。 本文采用光谱相异性指数(spectral dissimilarity index, BC)和光谱欧氏距离指数(euclidean distances, ED)定量估算两个样方之间的光谱差异。 计算公式如式(4)
式(4)中, BC表示两个样方间光谱的Bray-Curtis相似性, xis和xjs分别表示s波段在i和j样方的相对反射率, n表示波段数, BC值介于0~1之间, 0表示两个样方光谱组成完全一致, 1表示两个样方无共同光谱组成。
ED计算公式为式(5)
其中, xis和xjs与式(4)含义相同。
不同植物物种具有不同的生理生态特征, 在冠层上表现为不同的光谱反射特征[7, 8, 9, 10]。 不同植物物种的特征波段不同, 这是样方间物种beta多样性的一个有力指标。 采用已发表文献[7, 8, 9, 10, 11, 12]的物种特征波段作为光谱指数分析的波段(表1)。 对筛选的特征波段的相对反射率及其一阶导数求BC和ED值, 共获得164个高光谱指数, 用于估算beta多样性。
| 表1 用于估算beta多样性的植物生物学特征波段 Table 1 The characterized wavebands of plant biological traits used in the present study for extracting plant beta-diversity information |
对调查的270个样方, 随机分为两个数据集: 训练数据集(165个)和验证数据集(105个)。 基于1.3~1.5步骤计算高光谱指数。 通过相关性分析、 光谱指数稳定性和验证结果评估164个光谱指数。 筛选标准如下: (1)最优光谱指数与beta多样性指数的相关系数应显著; (2)筛选出来的光谱指数在不同群落条件下, 包括不同的盖度(≤ 25%, 26%~40%, ≥ 40%)和不同的物种丰富度(≤ 4, 5~9, ≥ 10)的估算精度应当保持稳定, (3)在验证数据集表现出显著相关性。 将估算出来的beta多样性指数值与实测的beta多样性指数值进行拟合, 具有显著相关性、 且决定系数较高的光谱指数将作为最优光谱指数。
164个高光谱指数与beta多样性指数相关性的结果表明, 物种BC值与高光谱指数呈现显著的相关系数最多, 物种Jaccard 和Sö rensen指数具有相同的显著性相关系数。 所选的特征波段中仅有部分与beta多样性显著相关。 420~480和490~570 nm的ED指数与物种BC指数、 Jaccard和Sö rensen指数显著相关。 与物种beta多样性指数相关性最多的指数将被选择, 通过相关性分析, 选择16个高光谱指数做进一步分析。
高光谱指数在不同群落条件下的稳定性, 是其应用潜力的一个重要评价依据。 本研究采用不同群落盖度和不同物种丰富度作为测试条件, 分析选择的16个高光谱指数与物种beta多样性的相关系数(表2)。 在不同群落条件下均与物种beta多样性显著的高光谱指数将作为潜在的最优指数。 经过比较, SIFD(400~1 000), EDFD(400~1 000)和SImeanFD(760~800)选为潜在最优指数。
| 表2 不同群落盖度和物种丰富度条件下高光谱指数与物种beta多样性之间的关系 Table 2 The Pearson coefficients between hyperspectral indices and species beta-diversity across different conditions |
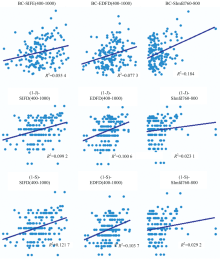
应用筛选的三个高光谱指数估算物种beta多样性, 将估算值与测试值进行回归拟合。 结果发现(图1), 估算值与实测值之间的回归系数达到统计学显著性(R2=0.02~0.2; p< 0.05), 表明筛选出来的高光谱指数基本符合要求。
 | 图1 实测值(y轴)与估算值(x轴)的散点图及其相关性(n=105)Fig.1 The scatter plots and their correlation coefficients (p< 0.01) between species beta diversity (y-axis) and hyperspectral beta diversity (x-axis) (n=105) |
因为研究地点在浑善达克沙地草地, 植物样方具有不同的盖度。 本文进一步分析了不同盖度对植物高光谱数据特征的影响(图2)。 图2表明, 即使是具有相同物种组成和丰富度的样方, 其样方相对反射率及其一阶导数表现不同。 这说明, 具有相同物种beta多样性的样方之间的光谱指数也可能不同。 这种影响降低了高光谱指数的估算精度。
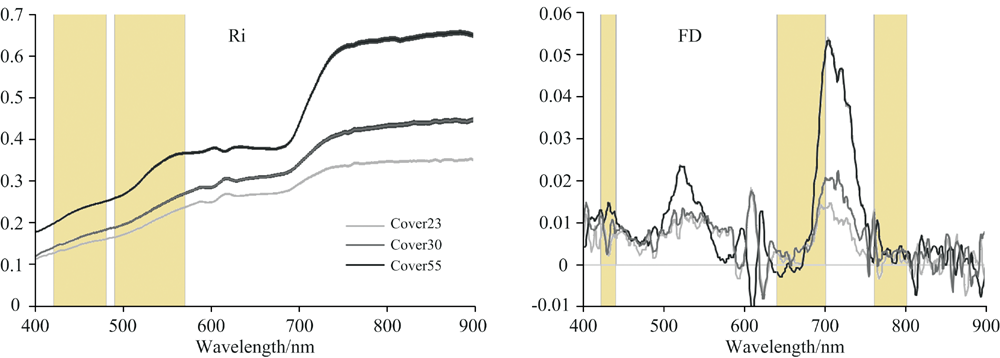 | 图2 具有相同物种组成但是不同多度的样方间的光谱反射率(Ri)和一阶导数(FD)曲线 注: 图中Cover23, 30, 55分别表示样方盖度为23%, 30%和55%; 图中阴影部分表示特征波段Fig.2 Plots with same species, richness and different abundance showing different reflectance (Ri) and (FD) curves under coverage 23%, 30% and 55% The gray columns indicate sensitive bands contained in proposed hyperspectral indices |
本研究表明, 可见光波段420~480, 490~570 nm和近红外760~800 nm波段与物种beta多样性指数显著相关。 这些波段也被认为是区分植物物种的特征波段。 经过三个步骤的筛选, 高光谱指数BCFD(400~1 000), EDFD(400~1 000)和BCmeanFD(760~800)能够估算植物物种beta多样性(R2=0.02~0.2; p< 0.05), 展现了高光谱数据估算物种beta多样性的潜力。
特征波段(420~480, 490~570和760~800 nm)中的420~480和490~570 nm位于叶绿体吸收高峰, 此波段范围的特征参数也能反映物种的差异。 760~800 nm位于植物细胞结构反射高峰区, 也经常用于进行植物物种区分。
虽然筛选出来的特征波段和高光谱指数与物种beta多样性之间密切相关, 但是这种关系还受到群落盖度的影响。 可见光和近红外波段处的光谱反射率随着植被盖度的增加而增加, 且近红外区增加更明显[11]。 本研究位于沙地草地, 植被覆盖度较低, 光谱指数受沙地背景的强烈影响。 总体上, 物种多样性指数与高光谱指数在中等的植被盖度(26%~40%)条件下相关性更高和稳定。 与光谱反射率相比, 一阶导数能够降低非植被的影响, 在不同群落盖度条件下估算物种beta多样性的表现较为稳定。 一阶导数能够反映相邻波段反射率之间的变异程度, 常被用于消除表面粗糙度和水分吸收引起的光谱变化[12]。 而且, 一些案例研究也表明, FD具有消除背景噪音和重叠光谱信号的作用[13]。 本文选择了特征波段的FD值作为高光谱指数, 获得了稳定的估算值, 也得益于FD值的作用。
本研究也表明, 不同的物种多样性指数与高光谱指数关系呈现一定差异。 其中, 物种BC指数与高光谱欧氏距离指数表现最为一致, 因为二者都考虑了组成元素数量的差异。 物种Jaccard和Sö rensen指数拟合效果较差, 其原因可能是二者都考虑了共同物种的数量, 但是在高光谱指数中难以找到相对应的变量。
研究选择的微尺度(0.8 m)也可能是改善估算精度的一个因素。 放牧干草地的案例研究表明, 随着空间尺度的增加(从3 m增加到6 m), 光谱相似性和物种相似性指数之间的相关性下降[14]。 其原因可能是随着空间尺度增加, 样方内的环境异质性(例如土壤理化性质和水分、 土壤类型、 枯落物和植被阴影)也会增加, 干扰群落的高光谱特征, 降低了物种光谱之间的差异性。
在浑善达克沙地草地, 采集高光谱数据和物种多样性数据, 采用相关性分析和特征波段筛选的方法, 计算了164个高光谱指数, 通过比较不同群落盖度和物种丰富度条件下高光谱指数估算精度的稳定性, 经过训练样方数据集和验证样方数据集的测试和验证, 筛选了精度较高的高光谱指数, 用于植物物种beta多样性估算。 这种方法能够减弱背景值的影响, 提高估算精度。 结果表明了高光谱遥感对于估算植物物种beta多样性的潜力。 随着机载、 星载高光谱传感器的广泛推广和应用, 高光谱数据估算物种beta多样性的研究和应用将会进一步发展。 未来的研究可以采用定量实验的方法, 比较不同植被类型、 不同时空尺度物种beta多样性的光谱模型, 发展更为稳定和适应性广的高光谱指数。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|