{kind=link}
{kind=link}
{kind=link}
{kind=link}
荧光分析技术在作物响应镉胁迫研究中的应用
[燕辉 , 李心平, 徐竹, 林国森]
, 李心平, 徐竹, 林国森]
, 李心平, 徐竹, 林国森]
|
|


作者简介: 燕 辉, 1984年生, 河南科技大学农业装备工程学院副教授 e-mail: yanhui@haust.edu.cn
为了揭示荧光分析技术在作物响应镉胁迫研究中的应用, 以大豆幼苗为材料, 对镉胁迫条件下叶片镉离子含量、 叶绿素荧光参数、 及荧光光响应曲线的变化进行了分析, 并进一步探讨了叶绿素荧光参数与叶片镉离子含量的关系。 结果表明: 在试验过程中, 伴随着镉胁迫时间的延长, 叶片镉离子含量呈现出逐步升高的趋势, 并在镉胁迫9 d后达到最大值。 镉胁迫1 d后, 大豆幼苗PSⅡ反应中心能够通过升高NPQ、 降低 ФPSⅡ与ETR来耗散过量激发能, 避免能量过激发对它的生理伤害, 进而维持 Fv/ Fm的稳定。 这表明此时的镉胁迫对大豆造成了动态光抑制。 镉胁迫6 d后, 虽然NPQ继续升高, 但光保护机制已不足以避免能量过激发造成的生理伤害, 大豆幼苗 Fv/ Fm呈现出降低趋势。 这表明此时大豆幼苗受到了慢光抑制的影响。 进一步对叶绿素荧光参数与叶片镉离子含量进行相关及回归分析发现: Fv/ Fm与镉离子含量呈非线性相关( R2=0.842, p<0.01), ΦPSⅡ( R2=0.959, p<0.01), ETR( R2=0.945, p<0.01)与镉离子含量呈线性负相关, 而NPQ( R2=0.959, p<0.01)与镉离子含量呈线性正相关。 这表明 Fv/ Fm能够在一定程度的镉胁迫下维持稳定, 而 ΦPSⅡ, ETR与NPQ均随着镉离子含量的升高而迅速发生变化。 对镉胁迫下叶绿素荧光的光响应曲线变化进行分析发现, 胁迫叶片 ФPSⅡ与NPQ光响应曲线呈现出与未胁迫叶片相似的变化趋势; 但在一定光强条件下, 胁迫叶片 ФPSⅡ较未胁迫叶片降低, 而NPQ较未胁迫叶片升高。 这表明镉胁迫导致叶片光化学活性降低, 激发能更多地以热的形式耗散。 以上研究证实, 荧光分析技术能够为深入研究作物响应镉胁迫的生理机制提供指导。
To reveal the application of fluorescence analysis technology on study of crop response to cadmium stress, the changes in cadmium concentration, chlorophyll fluorescence parameters, and fluorescence response curves of soybean seedlings under cadmium stress were analysed, and the relationship between chlorophyll fluorescence parameters and cadmium concentration in leaves was further assessed. The results showed that, with increased cadmium stress time, the concentration of cadmium in leaves increased gradually, reaching its maximum after 9 days of cadmium stress. After 1 day of cadmium stress, the PSⅡ reaction centre of soybean could dissipate excessive excitation energy by increasing non-photochemical quenching (NPQ) and decreasing actual efficiency of PSⅡ ( ФPSⅡ) and apparent electron transport rate (ETR). Thus the physiological damages caused by energy over-excitation in PSⅡ reaction centre were avoided and the stability of the maximal efficiency of PSⅡ ( Fv/ Fm) was maintained, indicating that the dynamic photo-inhibition in soybean caused by cadmium stress then occurred. After 6 days of cadmium stress, NPQ continued to increase, but the photo-protection mechanism was insufficient to avoid the physiological damage caused by energy over-excitation. Thus, Fv/ Fm of soybean seedlings showed a trend of decrease, indicating that soybean seedlings were then affected by chronic photo-inhibition. We next performed the correlation and regression analysis between chlorophyll fluorescence parameters and cadmium concentration. Fv/ Fm was non-linearly correlated with cadmium concentration ( R2=0.907, p<0.05), while ΦPSⅡ ( R2=0.959, p<0.01) and ETR ( R2=0.945, p<0.01) were negatively correlated with cadmium concentration. In addition, NPQ and cadmium concentration was positively correlated ( R2=0.959, p<0.01). These results indicated that Fv/ Fm could maintain stability under certain cadmium stress, while ΦPSⅡ, ETR and NPQ changed rapidly with increasing cadmium concentration. The changes in light response curves of chlorophyll fluorescence under cadmium stress were also analysed. It was found that the light response curves of ΦPSⅡ and NPQ in stressed leaves showed similar trends to those observed in non-stressed leaves. However, under specific light intensity, ΦPSⅡ in stressed leaves was lower than that of non-stressed leaves, while NPQ in stressed leaves was higher than that of non-stressed leaves, indicating that cadmium stress reduced the photochemistry activity of leaves, and caused more excitation energy dissipation as thermal. These studies confirmed that, fluorescence analysis technology could provide guidance for further study on the physiological mechanism of crop response to cadmium stress.
近年来, 随着工矿业“ 三废” 的过量排放、 化肥农药的滥施滥用、 及城市垃圾与交通污染的加剧, 土壤中的重金属污染呈现出不断加剧的趋势。 镉(Cd)在五大有毒元素(Cd, Hg, As, Cr和Pb)中被列为首位, 是一种毒性很强的重金属元素。 土壤中的镉离子极易被植物根系吸收, 并进一步被木质部蒸腾流运至冠层。 作为非必需元素, 冠层不断积累的镉离子不仅不能提供叶片必需的养分, 还会对叶片生长发育造成不利影响。 例如, 叶肉细胞中过量积累的镉离子会破坏膜蛋白氢硫键, 导致细胞膜、 细胞器膜与核膜透性增大, 进而造成细胞器与细胞核解体、 功能丧失[1]; 过量的镉离子还可能降低叶肉细胞中代谢酶活性、 限制叶片中营养元素吸收, 进而导致叶片发育不良、 失绿枯萎等[2]。 更为严重的是, 如果镉离子通过“ 植物— — 植食动物” 这一食物链进入人体、 并在人体内大量积累后, 必然会对人类的健康造成不利影响。
为了有效地获取植物响应镉胁迫的生物学信息, 就有必要依据植物的生理特性探求更为有效的分析手段。 通常情况下, 植物叶绿素分子吸收的光能分别用来: (1)驱动光化学反应; (2)以热能的形式耗散; (3)产生荧光, 且这三种能量释放方式之间相互竞争。 因此, 我们能够通过分析叶绿素荧光及热耗散的变化来探明植物光化学反应机制。 进一步研究证实, 叶绿素荧光分析能够灵敏、 快速、 无损地测定植物光系统II(PSII)反应中心活性, 进而获取光合过程中植物对光能的吸收、 传递、 利用与散失规律[3]。 因此, 与传统的利用植物叶片CO2释放量进行光合作用的研究相比, 它能够有效地反映植物光合作用的“ 内在性” 特点。 为探明荧光分析技术在作物重金属胁迫研究中应用的方法, 本研究以大豆幼苗为材料, 应用荧光分析技术, 探求镉胁迫下植物叶绿素荧光参数及其光响应曲线的变化规律, 以及荧光参数与镉离子含量的关系。 这将为在植物表型研究中探求植物逆境响应提供研究思路。
试验开始前, 首先将大豆播种于已填充基质的穴盘中。 待大豆出芽后, 选取生长良好且长势均匀的大豆幼苗, 分别移入统一规格的试验盆。 盆中填装等量基质, 且在整个试验期间确保水分充分供应, 以维持大豆幼苗正常生长。 待大豆幼苗生长至一定阶段后, 开始进行试验处理。 在试验过程中, 采用100 mL分别添加了0 μ mol· L-1(对照)与200 μ mol· L-1 CdSO4的溶液对大豆幼苗进行镉胁迫处理, 每种处理设置4个重复。 在镉胁迫1, 3, 6与9 d后分别测定对照与胁迫大豆幼苗的叶片镉含量与叶绿素荧光参数, 并对叶绿素荧光参数与叶片镉含量进行相关及回归分析。 同时, 在镉胁迫5 d后测定大豆幼苗叶片的荧光光响应曲线。
1.2.1 叶绿素荧光参数采集
镉胁迫1, 3, 6与9 d后, 采用Imagine-PAM调制叶绿素荧光仪(IMAGING-PAM, Walz GmbH, Germany)进行大豆幼苗叶片叶绿素荧光参数的测定。 在对叶片进行30 min暗适应之后, 首先采用光强小于 0.1 μ mol· m-2· s-1的一束弱光对叶片进行照射, 获取初始荧光(Fo)数据; 随后采用5 000 μ mol· m-2· s-1的饱和脉冲光进行0.8 s照射, 获取暗适应下最大荧光(Fm)数据; 再采用光化光进行一段时间的连续照射, 获取稳态荧光(Fs)数据; 最后采用饱和脉冲光对叶片进行照射, 获取光适应下最大荧光(F'm)数据; 光适应下初始荧光F'o=Fo/(Fv/Fm+Fo/F'm)数据可由荧光仪计算得到。
最大光化学效率为
Fv/Fm=(Fm-Fo)/Fm
实际光化学效率为
Φ PSⅡ = (F'm-Fs)/F'm
非光化学猝灭系数为
NPQ=(Fm-F'm)/F'm
电子传递效率为
镉胁迫5 d后, 采用Imagine-PAM调制叶绿素荧光仪进行大豆幼苗叶片荧光光响应曲线的测定。 测定时设定的光合有效辐射强度梯度为30, 60, 120, 250和500 μ mol· m-2· s-1, 测定的荧光参数包括实际光化学效率与非光化学猝灭系数等。
1.2.2 叶片镉含量测定
完成荧光测定后, 收获大豆幼苗叶片, 用液氮研成粉末。 称取一定量的粉末置于三角瓶中, 再向三角瓶分别加入10 mL硝酸与1 mL高氯酸。 次日, 将静置过夜后的三角瓶置于温度为100 ℃左右的电热板上消煮至0.5 mL左右。 随后, 将消煮液定容于容量瓶(25 mL), 并使用原子吸收光谱仪测定镉离子含量。
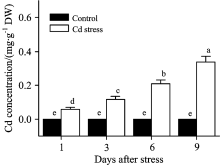
当植物根系暴露于镉离子环境时, 镉离子会通过主动或被动吸收的方式进入根细胞; 进入根区的镉离子会进一步在木质部蒸腾流的作用下向上运输至冠层, 并在叶肉细胞大量积累。 本研究亦发现, 伴随着胁迫时间的延长, 大豆叶片镉离子含量呈现出升高趋势, 且在镉胁迫9 d后达到最大值(图1)。 镉离子是非必需元素, 其大量积累不仅不会为植物生长提供养分, 还会对叶肉细胞质膜及细胞器造成生理伤害, 进而对光合器官及其生理功能造成不利影响。
 | 图1 镉胁迫下大豆幼苗叶片镉含量变化Fig.1 The variation of Cd concentration in leaves of soybean seedlings under Cd stress |
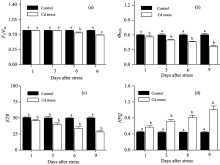
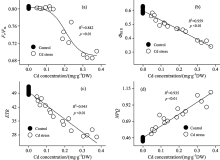
最大光化学效率(Fv/Fm)是反映光系统Ⅱ 反应中心原初光能转换效率或光系统Ⅱ 反应中心光化学潜在效率的物理量[4]。 对大多数植物来说, Fv/Fm是稳定的, 其值降低往往意味着植物遭受了环境胁迫或受到了生理伤害[5]。 对镉胁迫下大豆PSⅡ 反应中心Fv/Fm变化规律进行研究表明: 镉胁迫3 d后, Fv/Fm维持稳定, 这表明此时的镉胁迫没有对大豆PSⅡ 反应中心Fv/Fm造成影响。 但随着胁迫时间的延长, 镉胁迫6 d后, Fv/Fm较对照显著降低, 且随着镉浓度的增加降低幅度加大。 这表明此时镉胁迫对PSⅡ 反应中心造成了生理伤害。 整个试验中, 镉胁迫9 d条件下Fv/Fm值达到最低[图2(a)]。 回归分析表明, 大豆叶片Fv/Fm与镉离子含量呈非线性相关(R2=0.842, p< 0.01)[图3(a)]。 即Fv/Fm能够在一定程度的镉胁迫下维持稳定, 但随着镉浓度的进一步增大, Fv/Fm亦逐步降低。
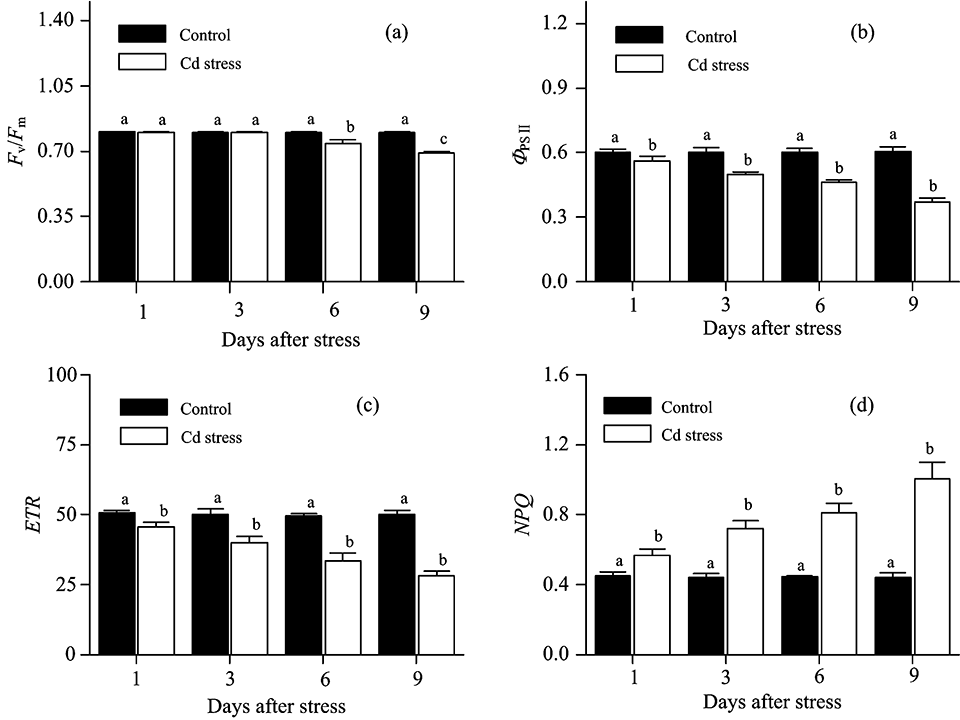 | 图2 镉胁迫下大豆幼苗叶片叶绿素荧光参数变化Fig.2 The variation of chlorophyll fluorescence in leaves of soybean seedlings under Cd stress |
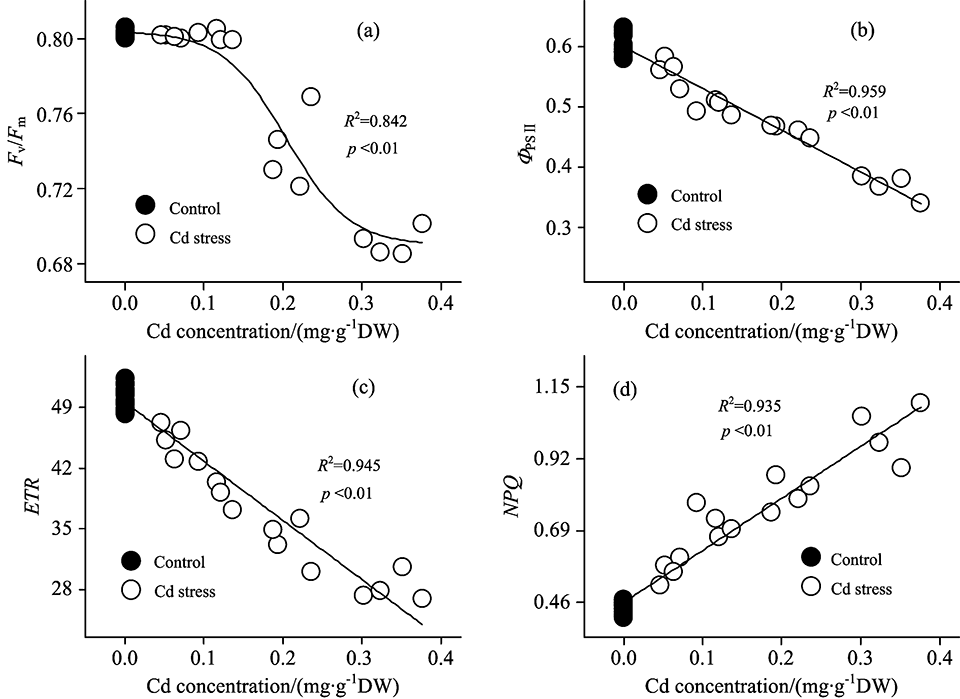 | 图3 大豆幼苗叶片叶绿素荧光与镉离子含量的相关及回归分析Fig.3 Relevant and regression analysis between chlorophyll fluorescence and Cd concentration in leaves of soybean seedlings |
实际光化学效率(Φ PSⅡ )指PSⅡ 反应中心在环境中部分关闭时的光化学效率[6]。 试验结果表明, 镉胁迫下Φ PSⅡ 呈现出与Fv/Fm不同的变化规律。 镉胁迫1 d后, 虽然Fv/Fm维持稳定, 但Φ PSⅡ 随镉含量的升高呈现出降低趋势。 这表明此时镉胁迫虽然没有对大豆光系统Ⅱ 反应中心造成生理伤害, 但已造成其实际光合效率降低。 伴随着胁迫时间的增加, 镉胁迫6 d对光系统Ⅱ 反应中心造成了生理伤害, 且Φ PSⅡ 随镉含量的增加降低更加显著[图2(b)]。 进一步的回归分析发现, 大豆叶片Φ PSⅡ 与镉离子含量呈负相关(R2=0.959, p< 0.01), 即随着叶片镉浓度的升高, Φ PSⅡ 逐步降低[图3(b)]。
电子传递速率(ETR)用Δ F/F'm× PFD× 0.5× 0.84(Δ F/F'm即Φ PSⅡ 表示PS Ⅱ 的实际量子产额、 PFD表示光合有效辐射、 系数0.84表示植物的经验性吸光系数、 系数0.5表示光能在植物两个不同光系统间平均分配)来计算[7], 表示光系统Ⅱ 反应中心的表观光合电子传递速率。 研究发现, 镉胁迫1 d后, ETR显著降低, 这意味着此时的胁迫程度已降低了光系统Ⅱ 表观光合电子传递速率; 随着胁迫时间的延长, 镉处理下光系统Ⅱ ETR下降幅度增大; 镉胁迫9 d后, ETR值下降至最低[图2(c)]。 回归分析亦表明, 大豆叶片ETR与镉离子含量呈负相关(R2=0.945, p< 0.01)[图3(c)]。
在PSⅡ 反应中心天线色素吸收的光能中, 部分能量不用于光合电子传递, 而是以热的形式耗散掉。 非化学猝灭系数(NPQ)是反映PSⅡ 反应中心热耗散能力大小的荧光参数[8]。 如果天线色素吸收的光能过量, 且不能及时有效地耗散, 将会对PSⅡ 反应中心造成伤害。 NPQ的值大, 说明天线色素吸收的光能更多地以热的形式耗散, 而用于光合电子传递和光化学反应的能量相对较少。 本研究发现, 镉胁迫1 d后, NPQ即随镉含量的增加呈现出升高趋势, 且镉胁迫3, 6与9 d后, NPQ均高于对照[图2(d)]。 表明镉胁迫条件下, PSⅡ 反应中心热耗散增加。 这与Ma等[9]对杉属植物的研究结论相一致。 大豆通过这种过量激发能耗散的方式, 可以有效地减轻镉胁迫下能量过剩对光合系统造成的生理伤害。 回归分析表明, 大豆叶片NPQ与镉离子含量呈正相关(R2=0.945, p< 0.01)。 即随着叶片镉浓度的增加, NPQ逐步升高[图3(d)]。
叶绿素荧光分析是研究植物光合特性的有效探针, 它有利于更深入地了解逆境条件下光合器官的状况[10]。 因此, 本研究对镉胁迫下大豆叶片叶绿素荧光参数变化规律进行了分析。 结果发现, 作为常用的叶绿素荧光参数, 光系统Ⅱ 反应中心Fv/Fm在一定程度的镉胁迫下能够维持稳定, 这与de Castro等[11]的研究结论一致。 随着镉浓度的增加, 100 mmol· L-1镉胁迫6 d后, 大豆幼苗Fv/Fm较对照显著降低。 Fv/Fm降低表明大豆遭受到了慢光抑制的生理伤害。 镉胁迫对大豆光系统Ⅱ 反应中心造成慢光抑制之前, 对其造成的动态光抑制就已经产生了, 主要表现在Φ PSⅡ 的降低。 Φ PSⅡ 的降低发生在Fv/Fm之前, 表明镉胁迫首先对植物造成了动态光抑制, 导致光系统Ⅱ 量子产额的降低。 随着镉胁迫程度的进一步增大, 光系统Ⅱ 反应中心的结构完整性受到伤害, 慢光抑制发生。
镉胁迫条件下, 伴随大豆光系统Ⅱ 反应中心Fv/Fm与Φ PSⅡ 的降低, ETR也呈现降低趋势。 这表明这些荧光参数之间存在相关性。 镉胁迫条件下, 光系统Ⅱ 反应中心光化学效率降低, 电子传递速率变慢。 而与之相反的是, 伴随着镉胁迫程度的增大, NPQ逐渐升高, 这确保捕光色素吸收的光能更多的以热的形式耗散掉而不是进行光合作用[12], 进而导致Φ PSⅡ 减量调节, 避免了原初电子受体的过度还原[13]。 但这种自我保护能力毕竟有限, 当镉胁迫程度继续加大且超出植物的承受能力(如100 mmol· L-1镉胁迫6 d)时, 导致发生慢光抑制及光系统Ⅱ 受损。
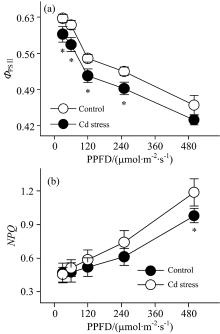
为了更有效地应用荧光分析技术评估作物响应重金属胁迫的生理机制, 本研究进一步对镉胁迫下大豆叶片叶绿素荧光光响应曲线变化进行了分析。 结果表明, 伴随着光强的增加, 大豆叶片实际光化学效率(Ф PSⅡ )呈现出逐步降低的趋势[图4(a)]; 而与Ф PSⅡ 不同, 大豆叶片非光化学猝灭(NPQ)随着光强增加逐步升高[图4(b)]。 这表明荧光参数的变化受光强的调控。 镉胁迫条件下, 胁迫叶片荧光参数光响应曲线的变化趋势与未胁迫叶片相似, 但一定光强条件下胁迫叶片荧光参数Ф PSⅡ 较未胁迫叶片降低[图4(a)]。 这应该与镉胁迫降低光系统Ⅱ 反应中心活性有关系[14]。 另一方面, 镉胁迫下反应中心Ф PSⅡ 的值下调能够降低光合电子传递速率, 进而减轻镉胁迫对反应中心的生理伤害。 而对镉胁迫下NPQ的光响应曲线进行研究发现, 一定光强条件下胁迫叶片的NPQ较未胁迫叶片升高[图4(b)]。 NPQ是一种光保护机制, 它能够将聚光色素吸收的光能更多地以热的形式耗散掉而不是进行碳同化[15], 进而减少能量过激发对作物的生理伤害。
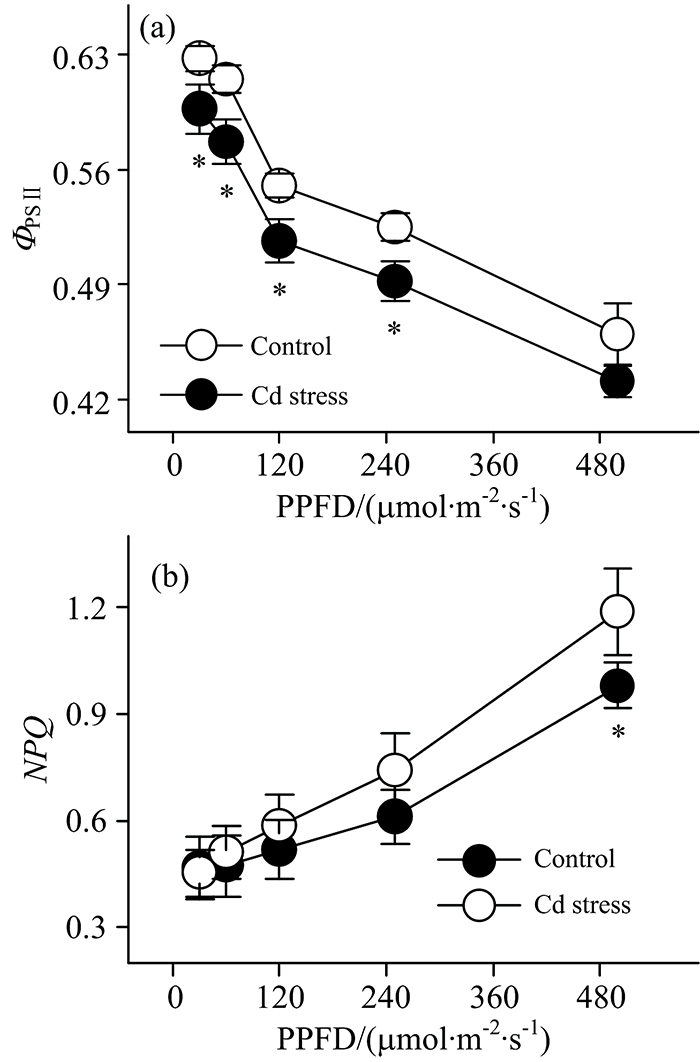 | 图4 镉胁迫下大豆幼苗叶片叶绿素荧光光响应曲线Fig.4 Light-response curves of chlorophyll fluorescence in leaves of soybean seedlings under Cd stress |
应用荧光分析技术探求作物响应镉胁迫的生理机制发现: 镉胁迫1 d即降低大豆幼苗PSⅡ 反应中心Ф PSⅡ 活性; 为了减少激发能过剩对PSⅡ 反应中心造成的生理伤害, 大豆幼苗通过升高NPQ耗散过剩激发能。 这虽然降低了PSⅡ 反应中心的光合电子传递速率, 但同时也避免了原初电子受体的过度还原, 维持了Fv/Fm的稳定。 此时镉胁迫对大豆造成了动态光抑制。 镉处理6 d后, 胁迫导致大豆幼苗PSⅡ 反应中心NPQ的进一步升高与Ф PSⅡ 、 ETR的进一步降低。 NPQ升高意味着光保护的增加, 然而, 此时的光保护已不足以避免能量过激发引起的伤害, Fv/Fm呈现出降低趋势。 这表明镉胁迫对PSⅡ 的慢光抑制已经产生。 胁迫叶片光响应曲线呈现出与未胁迫叶片相似的变化趋势; 但在一定光强条件下, 胁迫叶片Ф PSⅡ 较未胁迫叶片降低, 而NPQ较未胁迫叶片升高。 这表明胁迫导致叶片光化学活性降低, 吸收的光能更多地以热的形式耗散。 以上研究证实, 荧光分析技术能够为深入研究作物响应镉胁迫的生理机制提供理论指导。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|