


{kind=link}
{kind=link}
{kind=link}
{kind=link}
LED红蓝光连续光照及其光强对生菜生长及矿质元素吸收的影响
[查凌雁1, 2  , 张玉彬
, 张玉彬1, 2 , 李宗耕1, 2 , 刘文科1, 2, * ]
, 张玉彬]
|
|


作者简介: 查凌雁, 1991年生, 中国农业科学院农业环境与可持续发展研究所博士研究生 e-mail: zhaly2013@163.com
红光和蓝光是植物进行光合作用和光形态建成的主要光谱, 且红蓝LED是植物工厂光源的发展趋势。 因此为实现连续光照在植物工厂中的应用, 植物对红蓝光谱连续光照的响应特征及其机理亟待探究。 在环境可控的植物工厂内, 应用ICP-AES技术, 研究了红蓝光谱下连续光照及其光强对生菜生长和矿质元素吸收的影响。 该研究共包含两个试验, 试验一设置了常规光照(12 h/12 h)和连续光照(24 h/0 h)2个光照处理及5个生菜品种, 试验二设置了5个连续光照光强处理(80, 120, 160, 200和240 μmol·m-2·s-1)。 结果表明, 连续光照30 d内生菜的干重和干物质含量能够持续显著提高, 且表现为随连续光照光强增加而升高的趋势。 但地上部鲜重仅在连续光照的前15 d得到显著提高, 并随光强增加而升高。 而连续光照30天时, 连续光照对生菜地上部鲜重无显著促进作用, 甚至显著降低意大利生菜的地上部鲜重。 相比常规光照, 连续光照30 d显著降低了生菜的Ca, Mg, Fe, Mn, Cu和Zn含量, 但这些元素积累量均有所增加。 不同光强下, 生菜Ca, Fe, Cu和Zn的含量随光强增加而降低, 其中Ca和Fe积累量随光强增强而升高, Cu和Zn积累量不受光强影响。 Mg元素含量和积累量均随光强增强先升高后降低。 Mn元素含量受光强影响不显著, 但积累量随光强增大而升高。 此外, 连续光照30 d生菜叶片出现褪绿萎黄的伤害症状, 且伤害症状随光强增加而逐渐加重, 说明矿质元素含量的降低在一定程度上加剧了连续光照伤害。 综上所述, 15 d连续光照能够显著提高生菜产量。 全生长期(30 d)连续光照虽能促进生菜干物质累积, 但对产量无显著促进作用, 同时还会导致生菜矿质元素含量的降低及叶片伤害。 相对而言, 低光强(80和120 μmol·m-2·s-1)连续光照对生菜无明显伤害作用, 且矿质元素含量相对较高, 但不能提高产量。 研究结果表明短期红蓝光连续光照更适宜应用于在植物工厂生菜栽培, 能够获得相比能量投入更高的产量。 矿质元素含量的降低可能是长期连续光照伤害的机制之一。
Red light and blue light are the main spectra of photosynthesis and photomorphogenesis in plants, and also is a developing tendency of light source in plant factory. Therefore, to realize the application of continuous light in plant factory, the response characteristics and mechanism of plants to continuous light in red and blue spectra need to be explored. The effect of continuous light and its intensity on the growth and mineral elements absorption of lettuce was investigated by ICP-AES technology in an environmental-controlled plant factory. Five lettuce cultivars were cultivated under normal light(12 h/12 h) and continuous light (24 h/0 h) in experiment 1, and grown under continuous light with five light intensity(80, 120, 160, 200 and 240 μmol·m-2·s-1) in experiment 2. The results showed that dry weights and dry weight ratios of lettuce significantly increased during 30 days’ continuous light, and increased with light intensity. However, continuous light only enhanced fresh weight in first 15 days, which also increased with light intensity. 30 days’ continuous light had no positive effect on fresh weight, even significantly reduced the fresh weight of Yidali cultivar. Compared with normal light, 30 days’ continuous light significantly reduced the Ca, Mg, Fe, Mn, Cu, and Zn contents, but slightly or significantly enhanced the accumulation of these mineral elements. Under different light intensity, the contents of Ca, Fe, Cu, and Zn decreased with light intensity, the accumulation of Ca and Fe increased with light intensity, while that of Cu and Zn were not affected by light intensity. Content and accumulation of Mg increased first and then decreased with the increase of light intensity. The content of Mn element was not significantly affected by light intensity, but the accumulation increased with the increase of light intensity. In addition, 30 days’ continuous light induced severe leaf chlorosis and necrosis, and this negative effect aggravated with light intensity, which indicated that the decrease of mineral element content aggravated the injury of continuous light to some extent. In conclusion, 15 days’ continuous light could enhance lettuce yield remarkably. However, although 30 days’ continuous light improved the dry weight, it had no positive effect on yield and induced the decrease of mineral elements contents and leaf injury. Even though lettuce plants grown under continuous light with low light intensity (80 and 120 μmol·m-2·s-1) had no leaf injury and relative higher mineral elements contents, it had no positive effect on yield. This study indicated that short-term continuous light is more suitable for the cultivation of lettuce in the plant factory, and can obtain higher yield than energy input. The decrease of mineral element content may be one of the mechanisms of long-term continuous light injury.
光是植物生长发育最重要的环境因子之一, 它不仅可以作为能量来源和环境信号直接调控植物光合作用和形态建成, 而且还可通过光合作用影响植物某些酶的活性、 气孔的开闭、 蒸腾作用以及光合产物而间接调节植物矿质营养的吸收。 同时矿质元素也是光合作用中很多反应物质的必须组成元素以及多种酶的活化调节因子。 目前有关光质及光强对植物矿质元素吸收的影响已有较多研究[1]。 但关于光周期及光胁迫对矿质元素吸收影响的研究鲜有报道。 大量研究表明延长光周期是增加有效光照、 提高植物产量及品质的有效途径[2]。 在人工光植物工厂中, 利用人工光源可最大限度地延长光周期至连续光照。 前期研究证实适宜条件的连续光照能够显著提高植物产量和品质[3, 4]。 但也有研究表明, 连续光照会导致一些植物遭受胁迫伤害, 包括生物量降低、 植株早衰、 光合能力降低等[5, 6]。 其中, 最显著的症状是叶片褪绿萎黄。 有趣的是, 这种伤害症状和植物缺镁条件下表现出的萎黄病和坏死症非常相似。 并且这两种条件下的伤害症状都会随着光强及糖积累的增加而加重[7]。 说明连续光照下出现伤害的植株可能存在矿质元素的亏缺, 但目前有关连续光照对植物的养分吸收状况的影响尚无报导。 此外, 连续光照对植物的作用效果依赖于光强, 有研究表明生菜在弱光连续光照下能够正常生长, 且获得较高的生物量[8], 但在正常光强下则会受到胁迫伤害[9]。 因此, 探究连续光照及其光强对植物矿质元素吸收的影响, 有利于阐明连续光照伤害的机制并为连续光照在生产中的合理应用提供参考。
除光强外, 光谱组成也是影响连续光照效果的光环境因子。 但目前有关连续光照的研究大都采用全光谱的荧光灯或高压钠灯作为光源, 以致不同光谱成分连续光照的作用效果无法明确。 而且从植物光合有效辐射角度来看, 红蓝光谱是植物光合作用的主要作用光谱, 光合作用相对量子效率较高。 因此采用红蓝光谱进行连续光照有利于提高光能利用效率以及探究植物对连续光照的生理响应机理。 发光二极管(LED)因具有窄波段、 光谱光量可调控、 功耗低、 产热少等优势, 在植物对人工光源组合光谱的生长反应研究中广泛应用[10]。 已有研究证实LED红蓝组合光可以替代连续光谱栽培植物[11]。 生菜是一种被人们广泛食用的世界性蔬菜, 而且其体内所含的矿质元素是人体健康不可缺少的营养成分, 已有研究表明生菜矿质元素含量与光谱组成密切相关, 且LED红蓝光组合光下生菜矿质元素的含量显著高于荧光灯及自然光。 但红蓝组合光谱连续光照对其矿质元素含量的影响尚无报道。 因此, 本研究在环境可控的植物工厂内, 采用红蓝组合光谱LED为光源, 运用电感耦合等离子体原子发射光谱法(ICP-AES), 探究了红蓝光谱下连续光照及其光强对生菜生长及矿质元素吸收的影响, 以期为连续光照应用于植物工厂生菜生产提供策略。
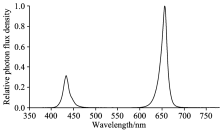
本研究包含两个试验, 均于中国农业环境与可持续发展研究所密闭植物工厂内完成。 以生菜(Lactuca sativa L.)为试验试材, 试验一供试品种包括: 美国大速生、 绿罗、 意大利耐抽薹、 汉斯206-紫悦和紫珊(下文中均用简称: 大速生、 绿罗、 意大利、 紫悦、 和紫珊)。 其中前3个品种为绿叶生菜, 后2个品种为紫叶生菜。 试验二依据试验一结果选择了对连续光照适应性相对较强的绿罗作为供试品种。 采用蛭石育苗, 待生菜幼苗长至两叶一心时, 移栽至水培槽中水培并开始进行光照处理。 营养液配方(mmol· L-1): 0.75 K2SO4, 0.5 KH2PO4, 0.1 KCl, 0.65 Mg SO4· 7H2O, 1.0× 10-3 H3BO3, 1.0× 10-3 MnSO4· H2O, 1.0× 10-4 CuSO4· 5H2O, 1.0× 10-3 ZnSO4· 7H2O, 5× 10-6(NH4)6Mo7O24· 4H2O, 0.1 EDTA-Fe, 4 Ca(NO3)2· 4H2O(pH: 5.90, EC: 1.24 mS· cm-1)。 试验期间植物工厂内昼夜温度为(25± 1) ℃/(22± 1) ℃, 湿度为60%70%, CO2浓度为外界大气CO2浓度。 选用LED红蓝光组合灯板(50 cm× 50 cm)进行光照处理, 红蓝光波峰分别为655和430 nm(图1)。 灯板悬挂水培槽上方45 cm处。
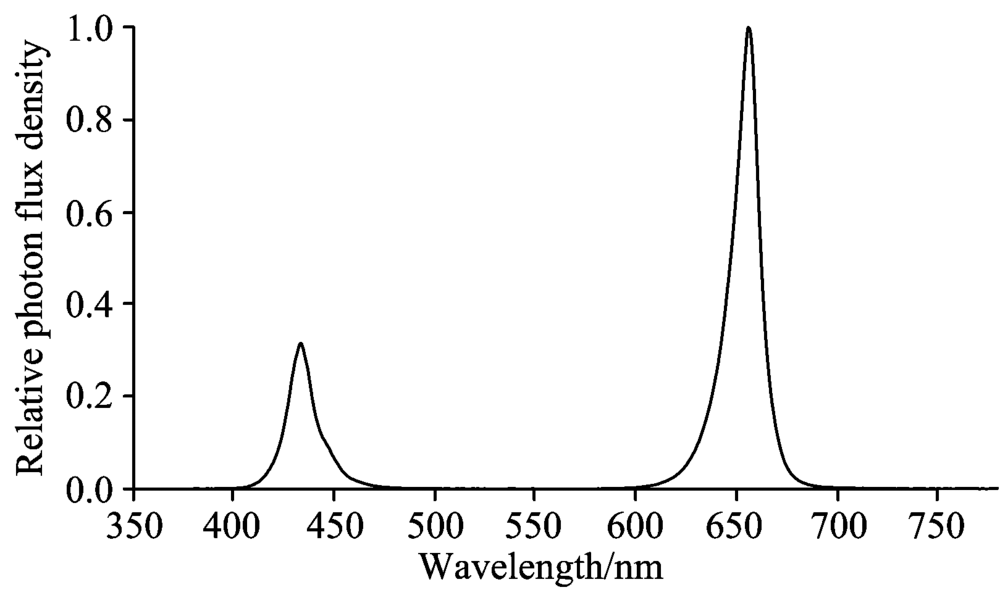 | 图1 LED红蓝光组合灯板光谱分布Fig.1 Spectra of red/blue LED lights |
试验一设置2个光周期处理: 常规光照(normal light, NL)和连续光照(continuous light, CL)。 光质和光强分别为3R: 1B, 240 μ mol· m-2· s-1。 试验二设置5个光照强度处理: 80, 120, 160, 200和240 μ mol· m-2· s-1。 光质和光周期分别为3R: 1B, 24 h/0 h(见表1)。 采用LI 1500辐射照度测量仪和LI-190R光合有效辐射传感器(美国LI-COR公司生产)测定栽培槽中心上方5 cm处光强, 并调节光质和光强至设定值。
| 表1 试验光照处理方案 Table 1 The details of light treatments |
两个试验均在移栽后的15和30 d取样测定生长指标, 每个处理随机选择长势具有代表性的 3 株生菜从茎基部分开, 称取地上部鲜重后将生菜105 ℃下杀青15 min, 以80 ℃烘干至恒重, 称取干重并计算干物质含量。 移栽30 d烘干的植物样品用组织研磨器研磨成粉状后精确称取1 g放入消煮管中, 加入高氯酸和浓硝酸的混合酸于180 ℃消煮, 冷却定容后用于测定矿质元素含量。 采用原子吸收分光光度计(ATC-006)和电感耦合等离子体质谱仪(ATC-155)测定矿质元素含量。
采用Microsoft Excel 2013软件对数据进行处理, 采用SPSS 16.0统计分析软件对数据进行差异显著性检验(Tukey法, α =0.05)。
由表2可见, 相比NL处理, CL处理15 d时5种生菜的地上部鲜重、 干重和干物质含量均显著提高。 除紫悦外, 其他四种生菜的干鲜重均提高了一倍以上。 5个品种中以大速生的生物量最高, 在NL和CL处理下均显著高于其他品种。 随着CL处理时长的增加, 5种生菜对CL的响应发生变化。 CL处理30 d时, 仅意大利生菜CL处理下地上部鲜重显著低于NL处理, 其他品种不同光照处理下地上部鲜重无显著差异, 但显著提高了地上部干重和干物质含量。 NL处理下5种生菜的干重和干物质含量均无显著差异, 但CL下5种生菜的干物质含量存在显著差异。 除30 d时的地上部鲜重外, 光照及品种对生菜鲜重、 干重和干物质含量的影响均达到极显著水平。 且光照是影响生菜生长的主要因素。
| 表2 LED红蓝光连续光照对5种生菜生物量及干物质含量的影响 Table 2 Effects of continuous light on the biomass and dry weight ratio of five lettuce cultivars |
表3和表4分别为连续光照下5种生菜矿质元素的含量和累积量, 可以看出, CL处理下5种生菜Ca, Mg, Fe, Mn, Cu和Zn的含量均低于NL处理, 其中Fe元素含量受CL影响最显著。 不同光照处理下, 5种生菜中以意大利生菜的整体矿质元素含量降低比率最高。 整体来说, 紫叶生菜的矿质元素含量相对绿叶生菜较高, 品种对Ca, Mg, Fe, Mn和Zn含量有极显著影响。 光照对6种矿质元素含量的影响均达到极显著水平, 且光照的F值明显高于品种及二者的交互作用, 说明光照是影响生菜矿质元素含量的主要因素。 除Fe和Zn外, CL处理下其余4种矿质元素的积累量均高于NL。 与NL相比, 大速生和绿罗的Fe含量在CL处理下显著升高, 但其他3种生菜的Fe含量却略有降低。 不同品种和光照处理间生菜的Cu积累量均无显著差异。
| 表3 LED红蓝光连续光照对5种生菜矿质元素含量的影响 Table 3 Effects of continuous light on the mineral elements contents of five lettuce cultivars |
| 表4 连续光照对5种生菜矿质元素累积量的影响 Table 4 Effects of continuous light on mineral elements accumulation of five lettuce cultivars |
不同时期, 生菜干鲜重及干物质含量随连续光照光强增加的变化趋势有所不同(图2)。 连续光照处理15 d时, 生菜的地上部干鲜重及干物质含量均随着光强增加而增加。 而在连续光照30 d时, 不同光强下生菜的地上部鲜重无显著差异。 同时生菜的干重和干物质含量维持着随光强增加而增加的趋势, 且不同光强处理间的差异显著性要高于15 d。
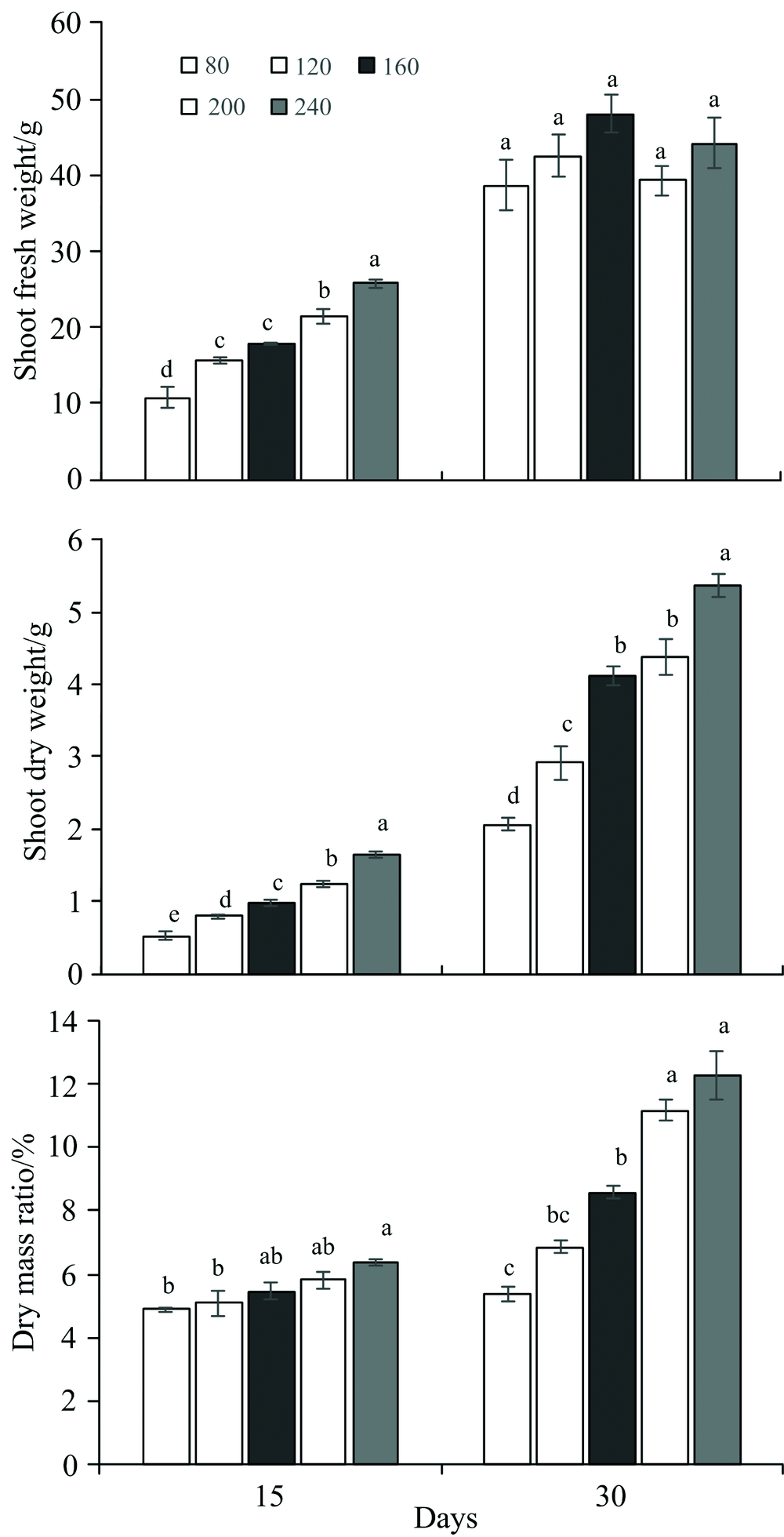 | 图2 不同光强下LED红蓝光连续光照对生菜生物量及干物质含量的影响Fig.2 Effects of continuous light with different light intensity on the biomass and dry ratio of lettuce |
如表5和表6所示, 连续光照处理30 d后绿罗生菜Ca元素含量和积累量分别表现为随光强增加而显著降低和升高。 Mg元素含量和积累量均表现为随着光强增加先增加后降低的趋势, 其中光强为160 μ mol· m-2· s-1时其含量和积累量均为最高。 连续光照光强对Fe和Mn元素含量无显著影响, 但其积累量随着连续光照光强的增加而增加。 Cu和Zn元素含量随连续光照光强增加而降低, 但其积累量不受连续光照光强影响。
| 表5 不同光强下LED红蓝光连续光照对生菜矿质元素含量的影响 Table 5 Effects of continuous light with different light intensity on mineral elements contents of lettuce |
| 表6 不同光强下LED红蓝光连续光照对生菜矿质元素积累量的影响 Table 6 Effects of continuous light with different light intensity on mineral elements accumulation of lettuce |
自然条件下光照时长是植物生物量增加的一个限制因子, 在园艺设施中可通过人工光源延长光照时长, 增加作物光合时间, 从而促进光合产物的积累, 提高产量[2, 3]。 连续光照最大程度地延长了光照时长, 理论上适宜条件的连续光照能够增加设施作物产量[12]。 本研究结果表明, 红蓝光连续光照15 d能够显著提高生菜的鲜重, 且鲜重随连续光照光强的增加而增加。 但红蓝光连续光照30 d对地上部鲜重无显著影响, 且不同光照强度之间也无显著差异。 干重对连续光照的响应与鲜重有所不同, 红蓝光连续光照15和30 d时, 生菜的地上部干重及干物质含量相比常规光照均显著增加, 且随连续光照强度增加也表现为显著增加趋势。 前人研究发现连续光照下, 多种植物干物质累积均显著增加[13]。 究其原因, 可能是因为连续光照增加了光合作用的时间, 同时避免了暗期呼吸作用的消耗, 从而增加了植株干物质的累积。 干物质比例的增加同时也意味着连续光照下植株含水率的降低, Arve等[14]研究发现番茄在连续光照下气孔的保水能力显著低于20 h光周期处理。 此外, 从生菜的外观形态可以看出红蓝光连续光照30 d时5种生菜均出现了显著的伤害症状, 主要表现为叶片斑点状萎黄褪绿, 幼叶叶尖坏死(图3)。 而且这种伤害症状在试验二中随光强增强而逐渐加重, 较低光强连续光照(80和120 μ mol· m-2· s-1)下虽无明显的伤害症状, 但生菜外观形态也表现出弱光下的生长特征, 如植株徒长、 叶片细长等(图4)。 针对连续光照产生伤害的原因, Velez-Ramirez[15]比较了日光光谱和红蓝光光谱连续光照对番茄生长的影响, 发现日光光谱下番茄的伤害症状更加显著, 并以此推断连续光照伤害并不是由光谱差异导致的。 本研究结果表明连续光照虽能够持续显著提高生菜的干重和干物质含量, 但全生长期连续光照对生菜鲜重无促进作用, 而且会导致生菜出现显著的伤害症状, 不利于其生长。 虽然降低光强能够缓解连续光照引起的伤害, 但对生物量无显著的提升效果, 且生菜外观形态异常。
 | 图3 LED红蓝光连续光照对生菜外观形态的影响Fig.3 Effects of continuous light on appearance and morphologyof five lettuce cultivars |
 | 图4 LED红蓝光连续光照光强对生菜外观形态的影响Fig.4 Effects of continuous light with different light intensity on appearance and morphology of lettuce |
植物对矿质元素的吸收主要是通过根表皮细胞的选择运输过程, 细胞膜上的载体蛋白是决定矿质元素吸收种类和数量的重要因素之一。 光可通过调节许多酶的活性而作用于多种蛋白质的合成进而影响矿质元素的吸收和运输[16]。 已有研究表明光谱成分对矿质元素吸收有显著影响, 相对而言, 有关光周期与植物矿质元素吸收间相互关系的研究较少。 前人研究发现随着光照时间的延长, 黄瓜对N, P和Ca特别是K矿质养分的吸收和积累大幅度增加[17]。 但本研究结果表明, 红蓝光连续光照30 d虽提高了生菜Ca, Mg, Fe, Mn, Cu和Zn的积累量, 却显著降低了这些元素的含量。 表明连续光照下生菜养分积累量的增加主要是因为植株干重的提高。 陈晓丽比较不同红蓝光比例对生菜矿质元素吸收的影响, 发现单一红色光谱下生菜Ca, Mg, Na, Fe, Mn, Zn, Cu和Mo含量均达最大, 但该光谱条件下生菜生物量最低, 导致一些元素的累积量反而最小。 说明光照可同时影响植物矿质元素含量和生物量, 从而影响其矿质元素积累。 本研究中连续光照光强越强, Ca, Fe, Cu和Zn元素含量越低。 整体而言, 生菜中矿质元素的含量与其伤害程度呈负相关的关系, 生菜的矿质元素含量越低, 其连续光照伤害症状越显著。 导致该现象的原因可能有以下几个方面。 首先长期连续光照会显著降低生菜的光合速率、 气孔导度和蒸腾速率[19], 蒸腾拉力是植物水分上升的主要动力, 矿质元素是以离子形式随水分经木质部向上运输, 其主要动力则是由叶片蒸腾作用产生的蒸腾拉力, 因而蒸腾速率的降低会影响矿质元素的吸收与运输。 其次, 连续光照会导致植物体内产生过量的活性氧(ROS), 有研究表明ROS的氧化损伤是导致植物缺Mg出现萎黄和坏死的原因[19], ROS能引起蛋白等生物大分子的损伤, 改变生物膜流动性、 离子运输等基本特性[19]。 关于光周期影响矿质元素吸收的机制, 还有研究认为光周期能够改变植物生长调节物的浓度[17], 而这些植物调节物能够影响植物根系对营养的吸收。 此外, 光周期还可能通过改变光敏色素系统来改变细胞膜对离子的渗透性[17]。 矿质元素是植物光合作用必需元素, 例如Mg是叶绿素的成分, 催化叶绿素合成的酶需要Fe2+激活, 而光合作用中水的光解需要Mn的参与。 因此, 连续光照下生菜体内矿质元素含量的降低会进一步加剧光合活性的降低和光合器官的伤害。 毫无疑问, 连续光照下植物必须吸收更多的矿质元素以维持正常的生长。 前人研究表明增加基质或营养液种的养分含量能够缓解连续光照对植物的伤害[13]。 因此, 在实际生产中可通过提高营养液浓度, 实现连续光照下生菜的生产。
全生长期(30 d)连续光照不适宜运用于生菜生产, 相对而言15 d的连续光照能够获得相比光能投入更高的生物量。 同时全生长期红蓝光连续光照导致会生菜矿质元素含量的显著降低, 并伴随着严重的叶片伤害症状。 连续光照引起的叶片伤害和矿质元素含量下降随其光强增加而加剧, 低光强连续光照下生菜虽无明显伤害症状且矿质元素含量相对较高, 但对产量无显著促进作用。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|