



{kind=link}
{kind=link}
{kind=link}
{kind=link}
三种典型荒漠植物生长期光谱特征变化分析
[加力戈1, 2  , 张勃
, 张勃1, * , 魏怀东1, 3 ]
, 张勃, 魏怀东|
|


作者简介: 加力戈, 1980年生, 西北师范大学地理与环境科学学院博士研究生, 讲师 e-mail: jialige@163.com
荒漠植物长势、 变化、 演替是反映荒漠地区生境状况的重要指标。 目前荒漠植物监测与光谱研究多基于定时采样数据, 波谱时序动态研究相对薄弱。 荒漠植物光谱因受时间尺度影响, 常引起辨识误差。 将荒漠植物中最具代表性的灌木——柽柳、 白刺、 梭梭作为样本, 旨在揭示三种荒漠植物光谱生长期变化规律及种间动态分异特征, 为荒漠植被空间遥感辨析奠定基础。 实验选取旺盛植株采集生长期内(5月—10月份)光谱数据, 对不同月份植物光谱曲线分析比较并剖析机理, 得出荒漠植物生长期光谱特征变化规律及其物候现象对应波谱表现。 结论指出: (1)三种荒漠植物反射率曲线总体特征均符合绿色植被波谱规律, 可观察到较明显的12峰谷分布, 红边斜率与面积从大到小分别为: 梭梭、 柽柳、 白刺。 其光谱曲线峰谷幅度值相对较小, 且变化较快, 红边参数表现活跃期分别为柽柳8月、 白刺10月、 梭梭9月。 (2)荒漠植物的光谱变化与植物本身物候特征、 气候变化植物响应密切相关。 光谱特征在可见光波段与营养期、 花期、 落叶期有一定响应关系; 近红外波段与结实期、 休眠期、 降雨情况相关; 短波红外波段与营养期、 落叶期、 降雨状况呈现关联性。 (3)7月份三种植物的生长状况差异光谱曲线表现为: 衰败植株地物光谱反射率可见光、 短波红外波段呈高反射, 近红外波段反射减弱, 趋近于土壤光谱反射率曲线。
The growth, change and succession of desert plants are significant indicators of the ecosystem in desert areas. At present, there are many studies on desert plant monitoring and desert plant spectrum based on fixed time sampling data, but few studies on a long time dynamic spectrum. The spectral discrimination of desert plants is due to the fact that it is affected by the time scale. In this paper, we selected plants which were representative of the desert shrubs— Tamarix ramosissima, Nitraria, Haloxylon ammodendron as samples, aiming at finding out the variation law and interspecific dynamic differentiation characteristics of the three desert plants, which was the basis for the ide.pngication of desert vegetation by spatial remote sensing. The spectral data of the vigorous plants were collected during the growing season (5~10 months) in the experiment, and the spectral curves of plants in deferent months were compared and analyzed. Results showed changes of the spectral characteristics and a corresponding phenology of desert plants in growing season. The conclusions are as follows. (1) The reflectivity curves of desert plants are consistent with the green vegetation spectrum. There are obvious peaks and valleys distribution. The slopes and areas of red-edge from large to small were as follows: Haloxylon ammodendron, Tamarix ramosissima, Nitraria. The amplitude of their spectral curve is relatively small, with rapid change. The active period of Red edge parameters is August for Tamarix, October for Nitraria, September for Haloxylon. (2) The spectral changes of desert plants are connected with the plant phenology and climate change. The spectral characteristics are related to the vegetative period, flowering period and deciduous period in the visible band, which are related to fruit period, dormancy period and rainfall in the near infrared band, and related to nutrition period, deciduous period, rainfall situation in the shortwave-infrared band. (3) In July, the spectral curves of three plants’ growth status showed that the spectral reflectance of decaying plant are higher than vigorous one in the visible band and mid-infrared band, while they are lower in the near-infrared band, which is close to the curve of soil reflectance.
地物光谱的测量与研究是遥感理论研究与应用分析的基石。 通过遥感实验测量的光谱元数据基础积累所建立的地物光谱知识库是面向众多行业科学的数据库。 光谱数据共享服务提供实时观测波谱数据并通过运行模型来外延观测数据。 植被光谱测量技术以对实验样本零破坏的优势, 迅速深入应用于植被特征研究领域, 尤其在植被资源匮乏、 生态环境恶劣地区更具现实意义[1]。
西北内陆干旱地区是荒漠植被分布最为集中普遍的典型区域, 也是沙漠化、 荒漠化重灾区。 荒漠植被光谱研究对植被理化性能分析及调查分类、 遥感反演等工作而言意义重大。 荒漠灌木— — 柽柳(Tamarix ramosissima)、 白刺(Nitraria)、 梭梭(Haloxylon ammodendron)在西北地区分布较广, 是最为典型的荒漠植被。 学者们对西北不同区域(甘肃安西[2]、 新疆准噶尔盆地[3, 4]、 甘肃民勤[5]、 新疆博州[6])荒漠植被反射光谱一般特征和红边参数特征等进行分析, 得出冠层光谱反射率差异较大且受土地背景的影响严重、 光谱特征相似但又区别于其他植被等结论。 经数十年的植物冠层反射光谱研究, 对一般的植被光谱特征变换分析方法由来已久[7]。 荒漠植被光谱特征相近, 遥感识别精度低, 常常用植物光谱特征指数及光谱曲线变换等方法深入讨论, 如红边参数分析、 一阶导数变换分析[8]、 包络线去除分析[9]、 倒数对数分析等。 与此同时针对各项指数分析植被元素含量(碳、 氮、 水分、 叶绿素、 纤维素)及生化特性的研究不断出现, 而荒漠植被的光谱指数与水分、 叶色素相关程度研究较为常见。
荒漠植物生长期(5月— 10月份)较短、 物候更迭时长短于普通中生植物, 光谱特征变化速度相对较快, 时间尺度影响更为明显, 光谱辨析与植被指数时相差异较大。 观测研究荒漠植被生长期时间序列光谱, 建立动态光谱特征数据库需求迫切。 因此生长期是绿色植被生物量聚增、 光合、 碳汇的主要阶段, 创造植被生产力的关键时期。 而荒漠植被光谱时间变化研究相对薄弱, 本工作尝试揭示三种荒漠植物光谱生长期变化规律, 对波谱特征区分及遥感匹配反演提供参考。
实验样本采集于甘肃省民勤县青土湖, 位于腾格里沙漠西北缘, 是石羊河尾闾, 海拔高度1 292~1 310 m, 年平均气温7.8 ℃, 10 ℃以上有效积温3 289.1 ℃; 年平均降水量89.8 mm, 降水多集中于7— 9月, 占全年降水总量的73%, 蒸发量达2 600 mm以上, 无霜期168 d。 研究区地理坐标为39° 04'14.8″N, 103° 36'56.5″E。 在1959年, 青土湖完全干涸后, 2010年通过渠道持续人工注水, 岸区生态因子好转, 周边主要荒漠植物群落结构得以恢复[10]。
在青土湖周边分别选取柽柳、 白刺、 梭梭作为样本, 2016年5月— 11月期间30 d间隔晴朗少云无风日采样6次, 观测时间均为北京时间上午10点至下午2点间, 并对环境参量进行详细记录。 在生长旺盛季节7月观测长势严重衰败植株作为对比样本。 见表1。
| 表1 光谱测量环境数据 Table 1 The environmental data of the spectroscopic observation process |
生长严重衰败样本获取于2016年7月2日上午、 长势: 严重衰退、 土质: 沙地、 经纬度: 39° 01'02″, 103° 18'32.8″、 海拔: 1 321.9 m。
测量方式采用FieldSpec4高分辨率地物光谱仪器, 测量高度选择20 cm, 试验中采用“ 独立工作模式” 特性允许使用者不需借助其他外界计算机等设备的情况下, 对350~2 500 nm光谱测量范围进行测量。 设备光谱分辨率(FWHM)≤ 3 nm: 350~1 000 nm, ≤ 16 nm: 1 000~1 900 nm, ≤ 14 nm: 1 900~2 500 nm辐射校正精度在400 nm: ± 5% 700 nm: ± 4% 1 000 nm: ± 5%, 符合野外测量需求视场角。
每种植物选取生长条件较好的10株, 各测量15次, 均值处理获得单次采样值与5次白板平均值比值的百分比算出单株该次反射率。 过程中确保所测量植株为单一植物样本。 每次测量新样本植被光谱时校正白板一次, 光谱仪探头垂直向下, 设置15° 探头视场角, 根据植被冠层的直径和高度确定探头距离冠层的高度, 保证视场可以覆盖冠层。
野外采样光谱数据因受到环境背景因素影响, 存在较多干扰信息, 需要对原始光谱数据进行鉴别筛选, 剔除无效值。 原始光谱数据读取转换采用ViewSpecPro6.2, 借助R语言平台对光谱曲线进行降噪及运算制图处理, 其中通过Technical Trading Rules(TTR)包内Simple Moving Averages(SMA)模型实现5 nm平均值滑动滤波方法。 对数据波动严重失真的1 350~1 450与1 800~1 950, 2 450~2 500 nm范围采用限幅50 nm滑动平均削弱干扰。
研究中通过一阶导数方法削弱背景噪声, 获取峰谷特征准确位置, 捕捉目标数据的变化并放大细小特征。 光谱数据受仪器设备光谱分辨率限制呈1 nm精度间隔离散数据信号。 因此通过R语言平台函数模型实现差分计算反射率一阶导数值。
研究对样本数据可视化处理, 先对其规律结合植物特性进行分析, 再将物候变化与光谱数据关键特征点时序变化相结合绘制成图。 文中植物物候资料参考常兆丰等学者民勤沙区主要植物的物候特征相关研究内容, 结合民勤沙产业生态园2016年物候现象记录开展讨论。
青土湖岸区属于典型干旱荒漠气候, 植物活动日数较短, 以灌木为主[11]。 为突出三种典型荒漠植物普遍代表性, 针对生长期六个阶段再均值处理后呈现光谱曲线[非光合作用植被(non-photosynthetic vegetation)样本光谱特征差异变化较大, 生长期均值综合特征曲线未包含在内]。 为便于分析描述, 对荒漠植物光谱曲线分别以12个特征点(可见光— — Visible(VIS)波段3个: 蓝谷、 绿峰、 红谷, 近红外— — Near-infrared(NIR)波段5个: NIR峰1、 NIR谷1、 NIR峰2、 NIR谷2、 NIR峰3, 短波红外— — Shortwave-infrared(SWIR)段4个: SWIR1谷、 SWIR1峰、 SWIR2谷、 SWIR2峰)进行定位, 如图1(a)。
3.1.1 三种荒漠植物的光谱特征
图1(a)中所示, 三种荒漠植被的反射率曲线总体特征在VIS(400~700 nm)范围内相近, 随波长增加反射率总体呈上升趋势, 梭梭的反射幅值高于白刺和柽柳, 其中青光波段(450~490 nm)480 nm处形成吸收率“ 蓝谷” 。 绿— 黄— 橙光波段(490~620 nm)处于低光合效率光谱带和弱活性带, 能量吸收较强, 并在550 nm处产生高反射“ 绿峰” 。 三种植物光谱均在红光波段(620~700 nm)680 nm附近形成明显的吸收谷, 称为“ 红谷” 。 在NIR波段(700~1 300 nm)680~750 nm处光谱曲线陡峭抬升形成绿色植物独有的强反射陡变特征“ 红边” 。 强烈抬升后在750~1 300 nm处, 出现高位波动持续的“ 高反射平台” 。 在SWIR-1(1 300~1 900 nm)和SWIR-2(1 900~2 500 nm)波段, 随波长增加反射率总体趋势下降。 其中高反射平台末端1 450, 1 950和2 450 nm呈现吸收谷, 进而在1 650和2 200 nm附近形成次递减的小反射峰。 三种植物光谱曲线趋势走向、 峰谷分布符合绿色植被光谱特征。
在NIR和SWIR-1的过渡区(1 400 nm附近)和SWIR-1和SWIR-2的过渡区(1 900 nm附近)受仪器自身噪声的影响及大气中水汽干扰, 波谱观测中出现强烈波动。 有研究表示环境介质中部分有机质造成强烈压抑吸收和反射时会产生敏感光谱— — “ 淬火效应” [9]。
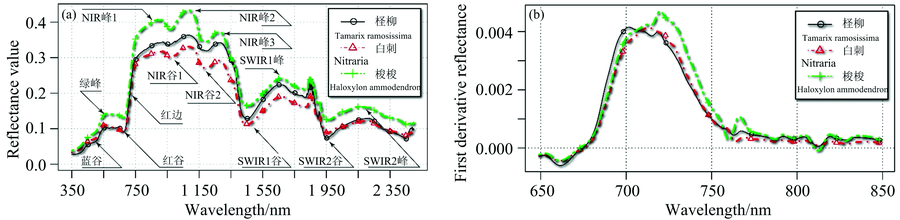 | 图1 (a)样本光谱反射曲线图; (b)红边反射波段一阶导数曲线图Fig.1 Spectral reflectance in (a) and its first derivative of Red edge in (b) |
3.1.2 三种荒漠植物光谱分析
图1(a)波谱曲线反射率阈值在3%~43%区间之内。 这与其他研究所获得的农作物反射率棉花2%~59%[4]、 水稻2%~49%[12], 油籽油菜2.5%~53%[13], 以及中生或湿生植被杨树4%~46%[14]等相比波幅范围较窄。 从大到小依次为梭梭(5%~43%)、 柽柳(4%~35%)、 白刺(4.5%~31%)。
在可见光波段峰谷分布与冠层叶绿素含量有关, 即取决于植物叶片中叶绿素含量、 冠层结构、 叶片面积、 枝叶结构情况。 蓝谷与红谷吸收带是植物低光合效率带, 大部分摄入能量由叶绿素所吸收。 由于叶绿素含量较高形成较高反射的绿峰, 可表征植物的健康程度。 三种样本“ 绿峰” 反射率在0.104 2~0.142 2区间, 从大到小依次为梭梭、 白刺、 柽柳, 这与直接对叶片目视观测绿度对比相一致。 红谷处梭梭反射率最高0.130 6, 柽柳略高于白刺分别为0.091 3和0.089 9。 由于光谱曲线分形维数可反映植被活跃状况, 在此通过峰谷值标准差反映光谱曲线变化程度。 在NIR高反射平台处, 柽柳、 白刺、 梭梭, 反射率均值为0.305 4, 0.270 5和0.350 4、 标准差为0.088, 0.080和0.096, 说明幅值及波动范围从大到小为梭梭、 柽柳、 白刺。 SWIR波段标准差柽柳最大0.062 2, 梭梭次之0.049 1, 白刺最小0.043 7。 其中SWIR1波段柽柳0.067 2、 白刺和梭梭为0.055 1和0.054 2, SWIR2波段柽柳0.039 5, 白刺0.022 9、 梭梭0.026 1。 说明SWIR段柽柳光谱曲线变化剧烈, 表现较强的生长活跃度。 白刺光谱曲线变化最为平缓。
在NIR/SWIR段, 三种植物光谱中对水分响应分别出现在970, 1 170, 1 450, 1 950, 2 450 nm处, 呈现吸收谷是荒漠植被储水能力的表现。 在残酷的自然环境选择过程中, 荒漠植被以顽强的适应性形成抗旱结构, 特殊的生理生态方式适应严酷的给养匮乏的环境条件。 枝叶结构产生适应性变化, 如叶面积速效退化减小蒸散发或肉汁茎叶储存水分构成与旱生植被的光谱特征有明显差异。
3.1.3 三种荒漠植物光谱红边一阶导数分析
红谷与近红外波段高反射平台之间形成的“ 红边” 是植物区别于其他物质的主要光谱特征。 高反射平台形成于叶绿素的多次散射, 可反映生物量、 叶面积指数等。 在此对生长期均值光谱红边一阶导数分析, 图1(b)。 红边导数图可见呈单峰分布, 且峰值位置相似, 分别为柽柳717 nm、 白刺718 nm、 梭梭719 nm, 由于对红光的强烈吸收红边位置均向长波方向靠近。 “ 红边” 面积受叶绿素含量、 冠层、 叶片结构等因素的共同影响[15], 从大到小依次为梭梭0.230 6、 柽柳0.210 7、 白刺0.200 5, 红边斜率为梭梭0.004 71、 柽柳0.004 15、 白刺0.004 10。
通过生长期光谱曲线定量对比评估, 开展三种退化植被动态研究。 三种植物不同月份生长期光谱曲线如图2。 对图2中三种荒漠植物光谱曲线分段分析, 并结合物候特征对三个波段光谱特征点变化对比分析如图3所示。 图3中对特征点值已归一化处理。
 | 图2 样本不同生长期光谱反射率曲线图(a): 柽柳; (b): 白刺; (c): 梭梭Fig.2 Spectral reflectance of samples at different growth stages(a): Tamarix ramasissma; (b): Nitraria; (c): Holoxylon ammodendrom |
 | 图3 三种荒漠植被特征点变化与物候对比分析图(a): 柽柳; (b): 白刺; (c): 梭梭Fig.3 Comparative analysis of desert plant’ s phenology and characteristic point changes(a): Tamarix ramasissma; (b): Nitraria; (c): Holoxylon ammodendrom |
3.2.1 柽柳生长期光谱逐月变化分析
结合图2(a)与图3(a)可知: 柽柳VIS波段, 5月份反射率幅度较高, 正至营养期(5月初), 介于展叶期(4月末)开花期(5月中旬至6月末)之间, 已有明显蓝谷、 绿峰。 整体反射高于其他月份, 低于10月。 6月份和7月份光谱曲线整体高于8月份和9月份, 且相互重合较多, 图2中难以区分。 6月份和7月份地球处于远日点是北半球光照最强的时间段, 植物荧光增强, 避免过度光照所造成的氧化伤害。 8月份叶绿素光合作用最佳, 光能量吸收最多, 也是柽柳果实成熟季节。 9月份绿峰略有削弱红谷吸收高于8月份, 进入营养期。 10月份反射率从青光波段开始明显高于与其他月份, 光合减少、 叶绿素下降, 进入落叶期, 植株老化趋势反射加剧。 NIR段反射率主要受叶结构影响。 5月份反射居中, 是花期的开始, 反射峰谷较为平均; 6月份反射率有所下降, 花期旺盛期。 7月份结实, 反射逐步增强高于5, 6月份; 8月份果实成熟, 反射率仅次于10月份。 NIR谷2、 峰3增幅缓慢。 10月份光谱曲线整体抬升与干物质含量增多有关, 可见光吸收减弱, 反射率达到最高。 SWIR段受叶含水量、 叶结构和干物质含量影响。 5月份两个峰向长波向依次下降。 SWIR1段, 6月反射率较5月有所下降, 7月略有上升, 8和9月份持续下降。 10月份反射率值明显升高, 峰值突出。 SWIR2段, 反射率值从大到小依次为10月、 5月、 6月和7月、 8月和9月。
3.2.2 白刺生长期光谱逐月变化分析
图2(b)与图3(b)可见, 白刺VIS部分光谱曲差异并不明显, 有较为明显蓝谷、 绿峰。 6, 7, 8和9月份阈值较低。 5月和10月反射率整体高于其他时段, 分别为白刺展叶期和落叶期, 紫光、 青光波段基本重合, 蓝谷长波方向10月底低于5月光能吸收。 绿光波段10月呈现较为明显的抬升延续至红谷。 6月份, 白刺花期绿峰向短波方向移动, 反射率高于区别并不明显的7, 8和9月份。 7月份反射率整体最低, 处于高温胁迫营养期。 8月份绿峰高于7和9月, 红谷最低, 叶绿素吸收活动呈现最强, 是白刺果实成熟期, 约持续20天。 9月由营养期转入叶变色期红谷抬升绿峰下降。 NIR、 SWIR段, 光谱曲线呈现5至8月下降, 8至10月抬升的规律, 8月份反射率最低。 从高到低依次为5, 10, 9, 6, 7和8月。 其中SWIR2谷处6月低于7月。
3.2.3 梭梭生长期光谱逐月变化分析
图2(c)与图3(c)显示梭梭样本VIS段, 5月份正至花期, 光谱曲线反射率高于其他5期样本, 绿峰未凸显。 6月至9月初为梭梭的较长营养期, 可见光吸收增强, 峰谷逐渐明显, 8月份蓝谷、 红谷吸收最强, 是光合作用逐渐加剧的过程。 9月中旬进入叶变色期、 月末果实成熟, 红谷明显抬升。 10月果实成熟及落叶期, 绿峰逐渐消退。 NIR段, 9月份果实成熟季节红边陡升, 峰谷明显, 反射最高。 NIR谷2、 峰3吸收较强。 5月曲线较为平缓, 低于9月。 6, 7和8月份是梭梭受高温胁迫的休眠期, 冠层叶倾角下垂, 反射率较低, 8月份梭梭生物活性缓慢复苏, NIR峰1抬升, 高于10月份。 峰2、 谷2、 峰3均低于10月, 由于雨水充足、 水分的吸收作用所致。 SWRI段, 季节分异规律性明显。 5至8月份下降, 8至10月份抬升。 其中9月份峰谷差异高于10月份。 反射率从大到小排列为5, 9, 10, 6, 7和8月份。
从时相变化来看, 柽柳8月份红边斜率与红边面积最大为0.006和0.267 9; 白刺的最大值出现于10月, 分别为0.005和0.227 0, 其次为8月0.004和0.201 9; 梭梭最大值出现于9月, 分别为0.007和0.337 0。 柽柳与梭梭红边参数高值基本出现在结实期与营养期, 白刺8月落叶期与营养期出现较高的红边参数, 可表示植物生长程度及生物量表现较为活跃。 三种荒漠植物红边参数在营养期均显示较高, SWRI波段营养期呈现吸收增强。
柽柳、 白刺、 梭梭是青土湖周边生态恢复期重要荒漠植物, 具备对多种不良生态条件适应能力, 是西北干旱地区水土保持改良树种。 由于大批量荒漠植被长势良莠不齐, 可借助光谱曲线反映三种荒漠植物健康状况, 如图4。
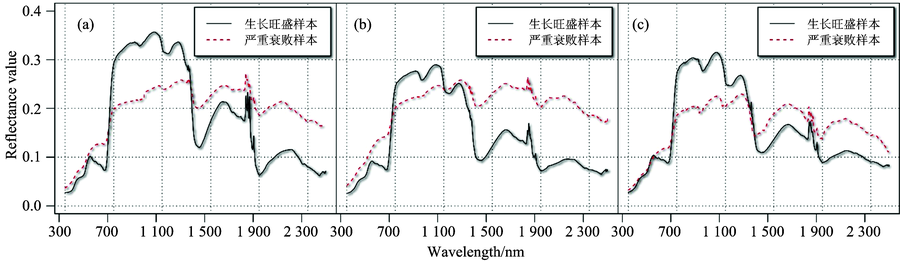 | 图4 七月生长旺盛期样本与衰败样本光谱曲线对比(a): 柽柳; (b): 白刺; (c): 梭梭Fig.4 Comparison of vigorous and poor growth sample in July(a): Tamarix ramasissma; (b): Nitraria; (c): Holoxylon ammodendrom |
三种植物7月严重衰败样本光谱曲线特征与健康样本相比, VIS波段呈现较高的反射率。 蓝谷略有抬升; 红谷处吸收减弱, 绿光波段反射率高于健康样本绿峰。 表现植物冠层色素偏红, 红光处呈现高反射。 可见光吸收减少, 光合作用减弱。 NIR峰1下降, 植物红边特征仍可见陡升, 逐渐丧失趋势明显。 高反射平台逐渐抬升, 峰谷差较小, 与生长较好样本曲线反射率差值在短波变大。 特征点削弱影响由两侧向红边位置增强。 SWIR段呈现较高的反射, 明显高于健康植物曲线, 向长波方向吸收逐步减小, 峰谷差距弱化。 与健康植物曲线NIR, SWIR1和SWIR2段的明显三阶梯式反射减弱表现相比, 衰败样本梯度效应明显减弱, 趋于平直。 但仍保持植物的光谱反射特征。
可见植物光谱反射率VIS和SWIR段高于健康植被, NIR段反之。 衰败程度加重, 更趋近于土壤光谱反射率曲线, 区别于健康样本曲线。 图4白刺7月样本光谱曲线衰败程度更为严重。
根据对柽柳、 白刺、 梭梭三种荒漠植被的生长期光谱数据的分析得出结论如下:
三种荒漠植物反射率曲线总体特征符合绿色植被光谱特征, 峰谷分布表现均可观察到较明显的12个特征点, 红边特征明显, 从总生长期长度观察, 红边斜率与面积从大到小分别为: 梭梭、 柽柳、 白刺。 因荒漠植物具备对严酷环境的适应能力, 光谱曲线峰谷幅度值相对较小, 且变化较快。 红边参数表现活跃期各不相同, 分别为柽柳8月、 白刺10月、 梭梭9月, 在营养期均呈现较高参数值。
从三种荒漠植物的光谱逐月变化分析, 可见曲线变化与植物本身物候特征联系紧密, 分布规律相互吻合。 可通过不同波段变化研究植物生长阶段性特点与气候变化植物响应, 通过物候观察与气象数据予以验证。 光谱特征在VIS段, 营养期、 花期、 落叶期有一定响应关系; NIR段, 与结实期、 休眠期、 降雨情况相关; SWIR段, 与营养期、 落叶期、 降雨状况呈现关联性。
荒漠地区生境严酷、 植被稀疏且生长状况差异明显。 在7月水热等要素最佳条件下, 三种植物的生长状况光谱曲线差异表现为: 衰败植株地物光谱反射率VIS和SWIR段呈高反射, NIR段反射减弱, 更趋近于土壤光谱反射率曲线。
本研究经野外调查获取大量的荒漠植物实地测量数据, 对典型荒漠植被光谱特性的深入挖掘构建荒漠植被时间序列光谱库积累资料。 结合物候观测资料分析光谱特征规律, 丰富荒漠植被光谱分析研究。 对荒漠植被的波谱区分及遥感反演奠定基础。 在后续研究中, 可通过荒漠植被敏感波段指数模型与传感器波段设计实现荒漠生态系统监测。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|